Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Рст - проксимальный извитой каналец. - дистальный извитой каналец. Рsт - проксимальный прямой каналец. Таl - дистальный прямой каналецСодержание книги
Поиск на нашем сайте
Транспорт ионов магния Почка в значительной мере принимает участие в балансе Mg2+, при этом выделение ионов контролируется гормонами. Паратиреоидный гормон снижает выведение Mg2+. В плазме крови находится 0,7-1,2 ммоль Mg2+ (частично связанного с белками), а в клубочковом фильтрате - около 0,5-0,9 ммоль. Экскретируемая фракция составляет <5%, максимально - 20%. Она возрастает при увеличении объема внеклеточной жидкости, гипермагнезиемии и гиперкальциемии, а также под воздействием диуретиков, влияющих на транспортные системы петли Генле, тогда как уменьшение объема внеклеточной жидкости, недостаток Ca2+ и Mg2+, а также гормоны действующие через cAMP (паратиреоидный гормон, кальцитонин и др.) снижают выведение Mg2+. В проксимальном канальце Mg2+ реабсорбируется значительно медленнее чем вода (и медленнее, чем Ca2+), поэтому концентрация Mg2+ в просвете канальца возрастает по отношению к концентрации (не связанного с белками) Mg2+ в плазме крови (с коэффициентом = 1,5), что вместе с положительным потенциалом жидкости в просвете канальца по отношению к плазме крови создает движущую силу для пассивной межклеточной реабсорбции Mg2+ в среднем и конечном отделах проксимального канальца (рис. 9-21 А). К концу проксимального канальца реабсорбируется около 15-20% Mg2+. Дистальный прямой каналец (толстая восходящая часть петли Генле) берет на себя большую часть реабсорбции Mg2+ (около 70%, это значение изменяется за счет регуляции) (межклеточный пассивный транспорт) (рис. 9-21 Б). Поскольку в этом отделе канальца трансэпителиальный потенциал тесно связан с происходящей там реабсорбцией NaCl, то изменения этой реабсорбции (например, за счет диуретиков, действующих на транспортные системы этого отдела, или в результате усиленного тока мочи) значительно влияют на реабсорбцию Mg2+. Наконец, дистальный извитой каналец реабсорбирует еще 2-8% профильтровавшегося количества Mg2+. Многие гормоны (паратиреоидный гормон, кальцитонин и др.) регулируют как трансклеточную реабсорбцию Mg2+ в дистальном извитом канальце, так и межклеточную реабсорбцию в толстом восходящем сегменте петли Генле. На межклеточный пассивный перенос может быть оказано влияние как за счет изменений положительного потенциала жидкости в просвете канальца (= движущая сила), так и за счет регуляции проницаемости плотных контактов. (В последнем процессе ключевую роль, по всей видимости, играет белок клаудин). Контроль за уровнем концентрации Ca2+ и Mg2+ в плазме крови осуществляется с помощью специального экстраклеточного сенсора: Ca2+/Mg2+-сенсора (Casr =Ca2+/Mg2+- sensing receptor), локализованного в гормональных железах, на базолатеральной мембране дистального прямого канальца и дистального извитого канальца. Рис. 9-21. Транспорт магния. Mg2+ реабсорбируются в проксимальном канальце - А (Ca2+ > Mg2+) и в дистальном прямом канальце - Б (толстой восходящей части петли Генле) (Mg2+ > Ca2+, см. также рис. 9-20) пассивно через плотные межклеточные контакты. Движущей силой этого процесса является положительный в просвете канальца трансклеточный потенциал, т.е. жидкость канальца заряжена положительно по отношению к плазме крови. В дистальном прямом канальце дополнительно осуществляется активная трансклеточная реабсорбция (Б). ICT - начальный отдел собирательного протока. CCT - корковый собирательный проток. 0MCD - наружный мозговой собирательный проток. IMCD - внутренний мозговой собирательный проток
Транспорт ионов калия Регуляция содержания К+ в организме осуществляется почками. Экскретируемая (выделяемая) фракция К+ составляет в среднем 5-15%, однако она изменяется в пределах между 1-3% при недостатке калия и 150-200% и при сильной гиперкалиемии. Поскольку реабсорбируемая фракция в проксимальном извитом канальце и в петле Генле (нисходящий тонкий сегмент петли - тонкий нисходящий отдел петли Генле, восходящий тонкий сегмент петли - тонкий восходящий отдел петли Генле, дистальный прямой каналец - толстая восходящая часть петли Генле) постоянна и составляет 85-90%, изменение выделения K+ обеспечивается связующими канальцами и собирательными протоками. Жизненно важно поддерживать концентрацию К+ в плазме крови в узких границах (обычно в пределах 4,1 +/- 0,6 моль/л). Поскольку почка в основном отвечает за выделение К+ в широком диапазоне, она должна реагировать на изменения концентрации К+ в крови (гипер- и гипокалиемия). Рассмотрим транспорт К+ в разных отделах нефрона. Экскретируемая фракция К+ составляет в среднем 5-15%, однако при недостатке К+ экскретируемая фракция К+ может быть уменьшена до значений 1-3%, при очень высоком потреблении или высвобождении К+ экскретируемая фракция может увеличиваться до 150-200%. К+ может полностью реабсорбироваться, а также полностью выводиться из организма с мочой. Эта приспособляемость реабсорбции К+ обеспечивается дистальным прямым канальцем (толстой восходящей части петли Генле) и собирательными протоками, поскольку, вне зависимости от потребностей К+-баланса, в проксимальных канальцах и в петле Генле (нисходящий тонкий сегмент петли - тонкий нисходящий отдел петли Генле, восходящий тонкий сегмент петли - тонкий восходящий отдел петли Генле, дистальный прямой каналец - толстая восходящая часть петли Генле) вместе взятых реабсорбируется постоянно 85-90% профильтровавшегося количества К+. Деполяризация апикальной мембраны главных клеток эпителия связующего отдела и собирательной трубочки стимулирует секрецию К+ этими клетками. Поскольку деполяризация мембраны зависит от регулируемой альдостероном электрогенной реабсорбции Na+, то секреция K+ тесно связана с реабсорбцией Na+ и зависит от концентрации альдостерона. Кроме того, сдвиг внутриклеточного значения pH в кислую сторону увеличивает проницаемость апикальной мембраны для К+ и следовательно секреция К+ увеличивается. За реабсорбцию K+ при его недостатке отвечают, по всей видимости, вставочные клетки эпителия связующих канальцев и собирательных протоков, в апикальной мембране которых, как и у обкладочных клеток желудка, локализована H+/К+-АТФаза, которая реабсорбирует К+ и секретирует в просвет собирательной трубочки ионы H+. Рис. 9-22. Транспорт K+ в корковых (кортикальных) нефронах. Невзирая ни на какие условия, в нефронах (А или Б), осуществляется реабсорбция и секреция K+. На рисунках представлены профильтровавшиеся фракции K+, реабсорбирующегося из просвета канальцев в интерстициальную жидкость (оранжевые заливки), и фракции K+, секретирующегося из интерстициальной жидкости в просвет канальца K+ (зеленые заливки). В проксимальных канальцах и в дистальном прямом канальце (толстой восходящей части петли Генле) постоянно реабсорбируется до 90% профильтровавшегося K+. Реабсорбция K+ осуществляется в этих отделах нефрона большей частью пассивно по межклеточному пути. Связующие канальцы, начальные отделы собирательных протоков и собирательные протоки имеют механизмы, обеспечивающие гомеостаз K+. При повышении содержания K+ (и потому высоком выделении альдостерона в кровь) главные клетки связующих канальцев и собирательные протоки могут секретировать большие количества K+, а при недостатке K+ (нет выделения альдостерона) вставочные клетки типа β этих отделов нефрона могут реабсорбировать K+
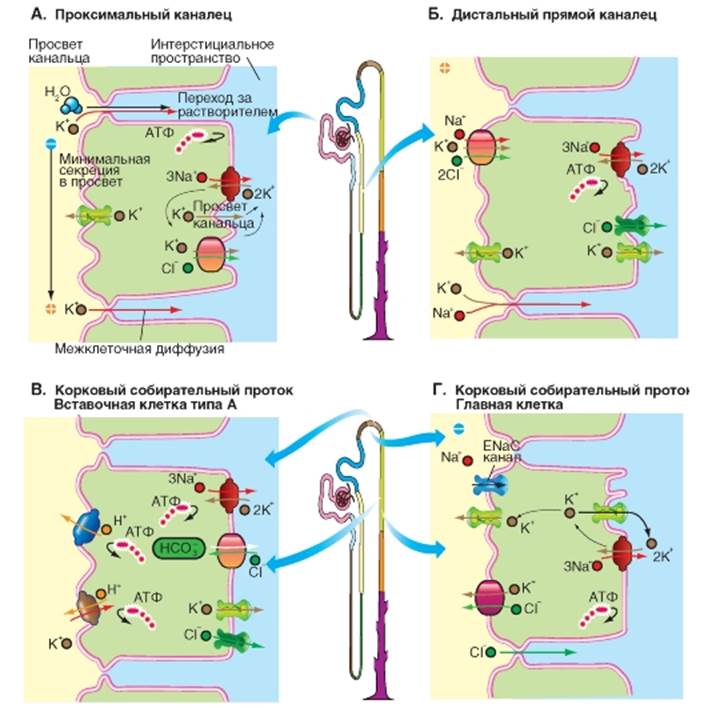
Клеточные механизмы транспорта K+ В проксимальных канальцах (рис. 9-23 А) в апикальной мембране с одной стороны происходит минимальная секреция K+ из клетки в просвет канальца через K+ -каналы, движущей силой чего является пассивная диффузия ионов. С другой стороны наблюдается реабсорбция в виде межклеточной диффузии или межклеточного перехода K+ вслед за водой. Это происходит большей частью за счет пассивного, межклеточного транспорта. В базолатеральной мембране Na+/K+ -АТФаза, потребляя АТФ, снижает внутриклеточную концентрацию Na+ и повышает внутриклеточную концентрацию K+ (первично-активный транспорт). K+ затем диффундирует из клетки через K+-каналы, локализованные в базолатеральной мембране (а не только в апикальной), что приводит к поляризации мембраны. Кроме того, на базолатеральной мембране расположен К+-С1--котранспортер, который переводит из клетки ионы K+ в интерстициальное пространство. В дистальном прямом канальце (толстая восходящая часть петли Генле) (рис. 9-23 Б) в апикальной мембране, обращенной в просвет канальца, имеется белок-переносчик (BSC1), который осуществляет одновременную реабсорбцию ионов Na+, K+ и 2C1-(механизм вторичного активного транспорта, котранспорт), при этом движущей силой является градиент Na+ опять создаваемый Na+/K+-ATФазой, расположенной на базолатеральной мембране (первичный активный транспорт). Однако на апикальной мембране наблюдается секреция K+ из клетки в просвет канальца через K+-каналы, движущей силой чего является пассивная диффузия ионов. Иначе говоря, поступающий в клетку с двух сторон K+ (переносчик BSC1 и Na+/K+-ATФаза) покидает клетку через опять же двусторонне локализованные K+-каналы. На базолатеральной мембране помимо Na+/K+-ATФазы и K+-каналов имеются C1- каналы, которые выводят из клетки в интерстициальное пространство избыток ионов C1-, закаченных переносчиком BSC1. Помимо всего прочего положительный в просвете канальца трансэпителиальный потенциал обеспечивает пассивную межклеточную реабсорбцию ионов Na+ и K+. Движущими силами этой межклеточной реабсорбции являются установившиеся в результате реабсорбции воды химические градиенты, положительный в просвете канальца трансэпителиальный потенциал - жидкость в просвете нефрона заряжена положительно по отношению к интерстициуму. Корковый собирательный проток (рис. 9-23 В, Г). Ультраструктурные особенности плотных контактов между клетками эпителия указывают на то, что в данном отделе нефрона мы имеем дело с относительно плотным эпителием. В этом отделе присутствуют, так называемые, вставочные клетки (intercalated cells), которые поодиночке разбросаны между многочисленными главными клетками (principal cells). Вставочные клетки находятся в корковом веществе почки и начальном отделе медуллярной собирательной трубочки. Они могут находиться в двух функциональных состояниях: их люминальная мембрана, обладающая большой поверхностью и снабженная микроворсинками, может быть либо выпячена в просвет канальца, (функциональный тип А), либо сокращена до небольшой поверхности (функциональный тип Б). Тип А секретирует ионы Н+ в просвет канальца, тогда как тип Б секретирует HCO3-. «Переключение» из А в Б подразумевает в том числе «обратное» встраивание транспортных протеинов для Н+ и HCO3-. Это демонстрирует, что регуляция транспортных свойств может включать в себя даже морфологическую перестройку клеток эпителия. В этой области плотные контакты собирательного протока сравнительно непроницаемы для катионов. Только C1- реабсорбируется в этом отделе нефрона межклеточным путем. В корковом собирательном протоке во вставочных клетках α -типа (рис. 9-23 В) в апикальной мембране осуществляется секреция в просвет канальца Н+ (Н+/K+-АТФаза) и переход во вставочную клетку К+. Кроме того в просвет канальца Н+ выкачивает Н+-АТФаза. В базолатеральной мембране находится белок-переносчик AE1 (антипорт HCO3-/C1-). Одновременно работает Na+/K+-ATФаза, закачивая во вставочную клетку К+. Однако эти ионы поступают в интерстициальное пространство по K+-каналам. В корковом собирательном протоке в главных клетках (рис. 9-23 Г) C1- реабсорбируется межклеточным путем. На апикальной мембране секреция К+ в просвет канальца зависит от реабсорбции Na+ по ENaC-каналам в клетку и потому от альдостерона и т.д. На люминальной мембране находятся K+-каналы и К+-С1-котранспортер, и обе эти системы секретируют K+ в просвет канальца. На базолатеральной мембране Na+/K+-ATФаза закачивает K+ в клетку, но его избыток уходит не только через K+-каналы люминальной мембраны, но и через K+-каналы базолатеральной мембраны. Рис. 9-23. Клеточные модели К+ транспорта. Рассматриваются четыре примера. А - проксимальный каналец. Б - дистальный прямой каналец (толстая восходящая часть петли Генле). В - вставочная клетка α-типа коркового собирательного протока. Г - главная клетка коркового собирательного протока
Функция концентрированной мочи В зависимости от количества воды, поступающей в организм, почки могут или выделять небольшое количество концентрированной мочи (0,3% клубочкового фильтрата; скорость образования мочи - 0,35 мл/мин; антидиурез (рис. 9-24 А); осмоляльность мочи около 1300 мосм/кг H2O) или большое количество гипотоничной мочи (20% клубочкового фильтрата; скорость образования мочи - 25 мл/мин; водный диурез (рис. 9-24 Б); 50 мосм/кг H2O). Для концентрирования мочи необходимы: (1) корково-медуллярный осмотический градиент, который устанавливается в результате реабсорбции NaCl в толстой восходящей части петли Генле и поддерживается противоточно-множительной системой мозгового вещества почек; (2) противоточный обмен в прямых сосудах (Vasa recta); (3) рециркуляция мочевины: медуллярная собирательная трубочка (проницаема для мочевины) - тонкая нисходящая часть петли Генле (проницаемы для мочевины) - толстый восходящий отдел петли Генле (непроницаем для мочевины) - дистальный извитой каналец (не-проницаем для мочевины) - медуллярная собирательная трубочка (проницаема для мочевины); (4) проницаемость собирательной трубочки для воды (вызванная антидиуретическим гормоном). Без антидиуретического гормона (АДГ) дистальный извитой каналец и собирательная трубочка непроницаемы для воды, поэтому гипотоничная моча, выходящая из толстого восходящего отдела петли Генле, за счет реабсорбции NaCl в последующих отделах нефрона становится еще более гипотоничной: водный диурез. Осмотический диурез вызывается нереабсорбируемыми профильтровавшимися веществами, тогда как диуретики тормозят реабсорбцию Na+ в различных отделах нефрона. Скорость образования мочи составляет у мужчин в среднем 1,35 и 1,15 л/сут у женщин, при этом наблюдается разброс индивидуальных значений в диапазоне от 0,5 до 2,0 л/сут. Если наблюдаемая скорость образования мочи соответствует значениям около нижней границы разброса (высококонцентрированная моча), то такое состояние называется антидиурезом, а при значениях около верхней границы разброса - диурезом. При количествах мочи больше 2,0 л/сут говорят о полиурии, при количествах меньше 0,5 или 1,0 л/сут об олигоурии и, соответственно, анурии. Кроме того, существует выраженная дневная периодичность, в ходе которой количество мочи рано утром на 40% ниже индивидуального среднесуточного значения, а в полдень - на 40% выше. Водный диурез. При переизбытке воды в организме прекращается выделение АДГ в кровь, и собирательная трубочка, как и дистальный извитой каналец, становятся непроницаемыми для воды, а проницаемость для мочевины уменьшается. В результате моча остается такой же гипотоничной, как и при переходе из петли Генле в дистальный каналец, а в дистальном канальце и собирательной трубочке она становится еще более гипотоничной за счет активной реабсорбции NaCl (минимально около 50 мосм/кг H2O). Подобное разбавление мочи обеспечивает выведение большого количества воды без одновременной потери NaCl и других веществ. При водном диурезе говорят о выведении «свободной» воды. Под этим подразумевается количество воды, которое может быть удалено из мочи, до тех пор пока ее осмоляльность не сравняется с осмоляльностью плазмы крови (Pосм = осмоляльность плазмы крови = 290 мосм/кг H2O). Относительная доля свободной воды в объеме мочи вычисляется из 1 - (U/P), при этом U - осмоляльность мочи. Если последняя составляет, например, 145 мосм/кг H2O, то доля свободной воды составляет 0,5 или 50%. Рис. 9-24. Осмолярность интерстициальной жидкости.
|
||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-20; просмотров: 578; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.143.7.53 (0.008 с.) |