Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Основные проводящие пути спинного мозгаСодержание книги
Поиск на нашем сайте
афферентной импульсации от рабочего органа управление им становится несовершенным; • способствует поддержанию тонуса ЦНС: при выключении афферентной импульсации уменьшается суммарная тоническая активность ЦНС; • участвует в процессах регуляции функций внутренних органов. Функции проводящих путей спинного мозга представлены в табл. 6.2. 6.12. Соматические рефлексы спинного мозга
Вегетативные рефлексы, в том числе осуществляемые с помощью спинного мозга, изучаются по главам, в разделах, посвященных регуляции функций органов и систем. Общая характеристика. Соматические рефлексы спинного мозга достаточно просты. По форме это в основном сгибательные и разгибательные рефлексы сегментарного характера. Надсегментарные рефлексы, наряду с сегментарными, осуществляются только с помощью шейного отдела. Рефлексы спинного мозга можно объединить в две группы по следующим признакам. Во- первых, спинальные рефлексы целесообразно объединить по органам (эффекторам рефлекса): рефлексы конечностей, брюшные, яичковый и анальный. Во-вторых, по рецепторам, раздражение которых вызывает рефлекс: проприоцептивные, висцероцептивные и кожные рефлексы. Последние являются защитными (см. табл. 6.2, 6.3). Рефлексы, возникающие с проприорецепторов, участвуют в формировании акта ходьбы и регуляции мышечного тонуса. Висцероцептивные рефлексы возникают с ин- терорецепторов (рецепторов внутренних органов) и проявляются в сокращениях мышц передней брюшной стенки, грудной клетки и разгибателей спины.
Защитные рефлексы, возникающие при раздражении кожи и тела, представлены в табл. 6.3 и 6.4. Рефлексы конечностей в клинической практике исследуют наиболее часто. Общепринятой классификации этих рефлексов не существует, их описывают без взаимной связи. Если за объединяющий признак рефлексов конечностей принять характер ответной реакции, то все их можно объединить в четыре группы. 1. Сгибательные рефлексы конечностей (фазные и тонические). Фазные рефлексы — это однократное сгибание конечности при однократном раздражении кожи или мышечных рецепторов. Рефлексы, возникающие с рецепторов кожи, являются полиси-
наптическими, они имеют защитное значение (см. рис. 6.11, А). Например, погружение в слабый раствор серной кислоты кончика лапки спинальной лягушки, подвешенной на крючке, или щипок кожи конечности пинцетом вызывают отдергивание конечности вследствие сгибания ее в коленном суставе, а при более сильном раздражении — ив тазобедренном суставе (сгибание конечности — избавление от раздражителя). Сгибательные (как и разгибательные) рефлексы, возникающие с мышечных рецепторов, являются моносинапти- ческими. Фазные проприоцептивные рефлексы участвуют в формировании акта ходьбы. По степени выраженности фазных сгибательных и разгибательных рефлексов определяют состояние возбудимости ЦНС и возможные ее нарушения. Поэтому в клинической практике исследуют несколько сгибательных фазных рефлексов. Ахиллов (пяточный) рефлекс— проприоцептивный, выражается в подошвенном сгибании стопы в результате сокращения трехглавой мышцы голени, возникает при ударе молоточком по ахиллову сухожилию; рефлекторная дуга замыкается на уровне крестцовых сегментов St—S2. Подошвенный ре.флекс — кожный, выражается в сгибании стопы и пальцев при штриховом раздражении подошвы; рефлекторная дуга замыкается также на уровне Sj-S2. Наиболее часто в клинической практике исследуют локтевой рефлекс — проприоцептивный, выражается в сгибании руки в локтевом суставе, возникает при ударе молоточком по сухожилию т. biceps brachii (при вызове рефлекса рука Должна быть слегка согнута в локтевом суставе); рефлекторная дуга замыкается в шейных сегментах спинного мозга (С5—С6). При возникновении фазных сгибательных рефлексов одновременно с возбуждением мотонейронов мышц- сгибателей происходит торможение мотонейронов мышц-разгибателей этой же конечности — иначе сгибание будет невозможным. Тонические сгибательные рефлексы конечностей участвуют в поддержании позы. Они возникают при длительном растяжении мышц и возбуждении мышечных рецепторов. Тоническое сокращение скелетных мышц является также фоновым для всех двигательных актов, осуществляемых с помощью фазных сокращений мышц. Изменение тонуса мышц конечностей свидетельствует о нарушениях в ЦНС. 2. Разгибательные рефлексы конечностей — моносинапти- ческие, возникают с проприорецепторов мышц-разгибателей. Фазные разгибательные рефлексы, в отличие от фазных сгибательных рефлексов, возникают в ответ на однократное раздражение только мышечных рецепторов, например при ударе по сухожилию четырехглавой мышцы ниже надколенника. При этом наблюдается коленный разгибательный рефлекс вследствие сокращения четырехглавой мышцы (мотонейроны мышц-сгибателей во время разгибательного рефлекса тормозятся с помощью постсинаптического реципрокного торможения посредством вставочных тормозных клеток Реншоу). Рефлекторная дуга коленного разгибательного рефлекса замыкается в поясничных сегментах (L2—L4). Фазные разгибательные рефлексы, как и сгибательные, участвуют в формировании акта ходьбы (перемещение тела в пространстве). Тонические разгибательные рефлексы — это длительное сокращение мышц-разгибателей при длительном их растяжении и возбуждении мышечных рецепторов. В положении стоя тоническое сокращение мышц-разгибате- лей предотвращает сгибание нижних конечностей и обеспечивает сохранение вертикальной естественной позы. Тоническое сокращение мышц спины удерживает туловище в вертикальном положении, обеспечивая осанку человека. Тонические рефлексы на растяжение мышц (сгибателей и разгибателей) называют также миотатическими.
Таким образом, главная роль тонических рефлексов — поддержание позы организма. 3. Ритмические рефлексы выражаются в многократном повторном сгибании и разгибании конечностей. Примером является рефлекс потирания: после смазывания раствором кислоты кожи бедра спинальная лягушка (лягушка, у которой удален головной мозг) многократно потирает этот участок — пытается освободиться от раздражителя. Слабое раздражение кожи боковой поверхности тела у собаки также вызывает ритмический рефлекс — почесывание этого участка задней конечностью. У спинального животного наблюдается также шагательный рефлекс. Шагательный рефлекс вызывается однократным раздражением кожи задней конечности, которое, в свою очередь, вызывает сгибание последней (см. рис. 6.11, А) и сопровождается дополнительным разгибанием противоположной задней конечности — перекрестный разгибательный рефлекс (см. рис. 6.11, Б). Затем согнутая конечность разгибается и опускается вниз, а разогнутая — сгибается и поднимается вверх, и так далее. Конечности не касаются опоры (собака зафиксирована в станке с помощью лямок), и без дополнительного раздражения они продолжают ритмично поочередно сгибаться и разгибаться — «шагать» в воздухе. Шагательный рефлекс (локомоция) может продолжаться часами, так как устранено тормозное влияние коры большого мозга. При неповрежденных сегментах, иннервирующих передние конечности (C5-Th2), и более длительном раздражении задней конечности в рефлекс вовлекаются также и передние конечности. Это наблюдение показывает наличие межсегментарной связи на уровне спинного мозга, играющей важную роль в ритмических движениях конечностей. Поочередное сокращение и расслабление мышц-сгибателей и мышц-разгибателей осуществляется в результате взаимодействия процессов возбуждения и торможения в соответствующих центрах спинного мозга под влиянием импульсов, поступающих в мозг от про- приорецепторов, что определяется их особым расположением относительно скелетной мышцы. Мышечные веретена (мышечные рецепторы, они первичные) расположены параллельно скелетной мышце — своими концами они крепятся к соединительнотканной оболочке (пе- римизию) пучка экстрафузальных мышечных волокон с помощью напоминающих сухожилия полосок соединительной ткани длиной 0,5—1 мм. В связи с этим, когда мышца расслабляется (удлиняется), растягиваются и мышечные рецепторы, что и ведет к их возбуждению (см. рис. 6.11, Л). Мышечный рецептор состоит из нескольких поперечнополосатых интрафузальных мышечных волокон (fusus— веретено), окруженных соединительнотканной капсулой и окончаниями афферентных волокон. Длина интрафузальных мышечных волокон составляет 4—7 мм, толщина — 15—30 мкм. Импульсы от мышечных рецепторов возбуждают нейроны своего центра и тормозят нейроны центра-антагониста. Ближе к концам мышечного веретена подходят двигательные нервные окончания, являющиеся аксонами у-мотонейронов спинного мозга. Их импульсация вызывает сокращение ин- трафузальных мышечных волокон (мышечные волокна рецептора), что ведет к возникновению РП, инициирующего посредством своего электрического поля возбуждение афферентного нервного волокна.
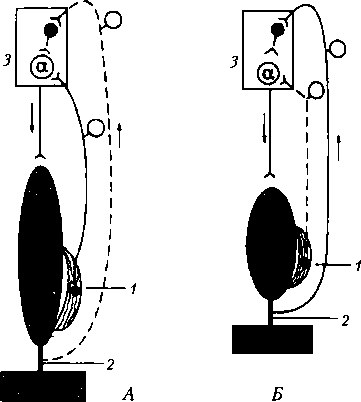
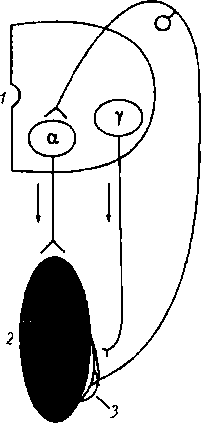
Сухожильные рецепторы (рецепторы Гольджи) крепятся относительно скелетной мышцы последовательно, что обеспечивает их раздражение при натяжении сухожилия (рис. 6.12). Поэтому сухожильные рецепторы посылают в мозг информацию о том, что мышца сокращена (напряжено и сухожилие), а мышечные рецепторы — что мышца расслаблена и удлинена. Импульсы от сухожильных рецепторов тормозят нейроны своего центра и возбуждают нейроны центра-антагониста (у мышц- сгибателей это возбуждение выражено слабее). Сухожильные рецепторы заключены в соединительнотканную капсулу и локализуются в сухожилиях скелетных мышц вблизи от сухожильно-мышечного соединения. Это тоже первичные рецепторы, они представляют собой безмиелиновые окончания толстого миелинового афферентного волокна — подойдя к капсуле рецепторов Гольджи, это волокно теряет миелиновую оболочку и делится на несколько окончаний, лишенных миелина. Механизм шагательного рефлекса. Когда мышца (сгибатель или разгибатель) расслаблена и удлинена, возбуждаются мышечные веретена, импульсы от них поступают к своим «-мотонейронам спинного мозга и возбуждают их
Рис. 6.12. Механизм возбуждения и торможения а-мотонейронов спинного мозга во время сокращения и расслабления скелетной мышцы: А — мышца растянута и расслаблена, возбуждаются мышечные рецепторы (/); Б — мышца сокращена (напряжена) и укорочена, возбуждаются сухожильные рецепторы (2); 1 — мышечные рецепторы (мышечные веретена); 2 — сухожилия мышцы и их рецепторы (рецепторы Гольджи); 3 — полусегмент спинного мозга. Афферентная импульсация: сплошная линия — выражена, пунктирная линия — отсутствует
(см. рис. 6.12). Затем а-мотонейроны посылают импульсы к этой же скелетной мышце, что ведет к ее сокращению. В результате сокращения мышцы возбуждение мышечных веретен прекращается или сильно ослабляется (они уже не растянуты), начинают возбуждаться сухожильные рецепторы, поскольку сухожилие натягивается (см. рис. 6.12, Б). Импульсы от сухожильных рецепторов поступают тоже, в первую очередь, в свой центр в спинном мозге, но к тормозным клеткам. Возбуждение этих клеток вызывает торможение а-мотонейронов этой же скелетной мышцы, вследствие чего она расслабляется. Однако ее расслабление (удлинение) ведет снова к возбуждению мышечных веретен и а-мотонейронов — мышца вновь сокращается. Вследствие ее сокращения снова возбуждаются сухожильные рецепторы и тормозные клетки в спинном мозге, что ведет к очередному расслаблению скелетной мышцы, и так далее. Мышца поочередно сокращается и расслабляется в результате поступления к ее мотонейронам импульсов от собственных рецепторов. Описанные процессы в равной степени относятся и к мышце-сгибателю, и к мышце-разгибателю, при этом расслабление скелетной мышцы запускает механизмы ее сокращения, а сокращение скелетной мышцы активирует механизмы, расслабляющие мышцу, что наблюдается при ходьбе и беге или любом повторяющемся действии.
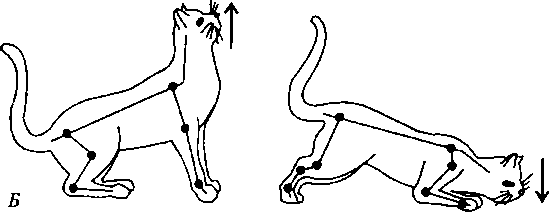
Таким образом, и мышцы-сгибатели, и мышцы-разгибатели рефлекторно сокращаются только с помощью мышечных рецепторов, а рефлекторно расслабляются только с помощью сухожильных рецепторов. Во время ходьбы в естественных условиях мышцы-сгибатели и мышцы-разгибатели одной и той же конечности должны сокращаться и расслабляться последовательно друг за другом, что достигается с помощью торможения центра-антагониста при возбуждении центра-агониста (см. рис. 6.12, А). Причем если на одной ноге сокращены сгибатели, на другой ноге сокращаются разгибатели, иначе нарушится поза. Это обеспечивается поступлением афферентных импульсов от мышечных и сухожильных рецепторов и поочередным возбуждением и торможением центров-сгибателей и центров-разгибателей (см. рис. 6.12, Б) — перекрестный разгибательный рефлекс. Шагательные координированные движения возможны в отсутствие обратной афферентации от проприорецепторов. Они осуществляются с помощью межсегментарных связей на уровне спинного мозга и генератора локомоции каждой конечности. О наличии межсегментарных связей на уровне спинного мозга свидетельствует также факт вовлечения в шагательный рефлекс всех четырех конечностей при достаточно длительном и сильном раздражении одной конечности. Однако у человека после разрыва спинного мозга локомоция не восстанавливается, но эти механизмы включаются вышележащими отделами ЦНС. 4. Рефлексы позы спинального организма (шейные рефлексы) заключаются в перераспределении тонуса мышц конечностей при изменении положения шеи относительно туловища. Они осуществляются с участием различных отделов ЦНС. На уровне спинного мозга замыкаются шейные по- зные рефлексы, наличие которых установил голландский физиолог Р. Магнус (1924) в опытах на кошке. Имеется две группы этих рефлексов. Первая группа шейных позныхрефлексов возникает при наклоне головы вперед или назад (рис. 6.13). При наклоне головы вниз (кпереди) увеличивается тонус мышц-сгибателей передних конечностей и тонус мышц-разгибателей задних конечностей, в результате чего передние конечности сгибаются, а задние — разгибаются. При наклоне головы вверх (кзади) возникают противоположные реакции: передние конечности разгибаются вследствие увеличения тонуса их мышц-разгибателей, а задние конечности сгибаются вследствие повышения тонуса их мышц-сгибателей. Эти рефлексы возникают с проприорецепторов мышц шеи и фасций, покрывающих шейный отдел позвоночника. В условиях
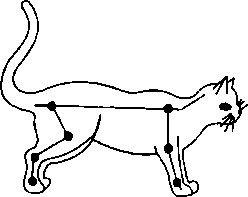
Вторая группа шейных позных рефлексов (тонических) возникает с шейных проприорецепторов при поворотах или наклонах головы вправо или влево (рис. 6.14). Рефлекс выражается в повышении тонуса мышц-разгибателей обеих конечностей на стороне, в которую повернута (наклонена) голова, и в повышении тонуса мышц-сгибателей на противоположной стороне. Рефлекс направлен на сохранение позы, которая может быть нарушена вследствие изменения положения центра тяжести после поворота (наклона) головы. Центр тяжести смещается в сторону поворота головы — именно на этой стороне по-
Рис. 6.14. Изменение тонуса мышц конечностей при наклоне головы вправо (Л) и влево (Б) (шейные тонические рефлексы) вышается тонус мышц-разгибателей обеих конечностей, что повышает устойчивость организма. Подобная ситуация наблюдается и у человека. 6.13. Тонус мышц у спинального организма Спинальный шок. В эксперименте тонус скелетных мышц исследуют после поперечной перерезки спинного мозга. Сразу после перерезки или повреждения спинного мозга у человека в случае травмы наблюдаются мышечная атония и отсутствие рефлексов (спинальный шок). Главной причиной спинального шока является выключение влияния вышележащих отделов ЦНС на спинной мозг. Об этом свидетельствует тот факт, что повторная перерезка спинного мозга ниже первой повторно шок не вызывает. Спинальный шок у лягушек длится несколько минут, у собак — несколько дней, у человека — около 2 мес. После исчезновения спинального шока тонус мышц, иннервируемых посредством сегментов спинного мозга, которые находятся ниже перерезки (повреждения), резко повышается. Гипертонус мышц спинального организма. Сгибательные и разгибательные рефлексы нижних конечностей у человека в случае повреждения спинного мозга усиливаются (рис. 6.15). Гипертонус имеет рефлекторную природу: деаф- ферентация, например, нижней конечности у лягушки ведет к исчезновению тонуса у нее (опыт Бронжеста). Однако произвольные движения невозможны! Афферентная импульсация от мышечных рецепторов по задним корешкам спинного мозга (см. рис. 6.15), которая ведет к возбуждению а-мотонейронов спинного мозга и сокращению скелетных мышц и развитию гипертону-
Рис. 6.15. Механизм регуляции тонуса мышц на уровне спинного мозга и развития гипертонуса мышц нижних конечностей в случае повреждения спинного мозга:
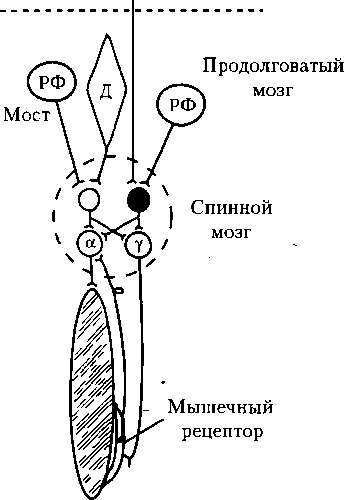
1 — полусегмент спинного мозга; 2 — скелетная мышца; 3 — мышечное веретено; а — мотонейрон, иннервирующий миоциты скелетной мышцы; у — мотонейрон, иннервирующий миоциты интрафузального мышечного волокна (пропри- орецептора) са, поддерживается двумя способами: 1) спонтанной активностью мышечных рецепторов, что обеспечивает поступление импульсов от них к а-мотонейро- нам, возбуждение последних и сокращение мышц; 2) спонтанной активностью ^-мотонейронов. Возбуждение у-мотонейронов вызывает возбуждение и сокращение иннервируемых ими интрафузальных мышечных волокон, в результате чего увеличивается натяжение мышечного рецептора, поскольку концы его зафиксированы на скелетной мышце. Это вызывает раздражение и возбуждение мышечных рецепторов, импульсы от которых поступают к а-мотонейронам и возбуждают их. В свою очередь, а-мотонейроны посылают импульсы к скелетной мышце и вызывают ее постоянное (тоническое) сокращение, вследствие чего и развивается гипертонус у спинального организма. Следует отметить, что в нормальных условиях деятельность спинного мозга контролируется вышележащими отделами ЦНС, нисходящие влияния которых являются преимущественно тормозными. 6.14. Двигательные системы ствола мозга Ствол мозга включает продолговатый мозг, мост и средний мозг, где локализуются ядра черепных нервов продолговатого мозга (IX-XII нервов), моста (V- VIII пары) и среднего мозга (III, IV пары), а также другие ядра, управляющие мускулатурой туловища и конечностей (см. п. 6.14). Функции отдельных ядер черепных нервов рассмотрим в восходящем ряду: продолговатый мозг, мост и средний мозг. Двигательное ядро добавочного нерва (XI, п. accessorius, продолговатый и спинной мозг), иннервируя грудиноключичную и трапециевидную мышцы, обеспечивает их сокращение и поднимание плечевого пояса кверху, лопаток — к позвоночнику, повороты и наклоны головы. Ядра языкоглоточного нерва (IX, п. glossopharyngeus, продолговатый мозг). Чувствительное ядро одиночного пути формирует все виды чувствительности слизистой оболочки глотки, задней трети языка, барабанной полости и каротидного тельца, участвует в рефлексах жевания, глотания, в секреторных и моторных пищеварительных рефлексах, в сосудистых и сердечных рефлексах (из каротидного тельца). Нижнее слюноотделительное (парасимпатическое) ядро стимулирует секрецию околоушной слюнной железы. Двойное (двигательное) ядро обеспечивает поднимание глотки и гортани, опускание мягкого нёба и надгортанника в глотательном рефлексе. Двигательное ядро подъязычного нерва (XII, п. hypoglos- sus, продолговатый мозг) иннервирует мышцы языка и обеспечивает его движение в процессе разговора, жевания, глотания, сосания. Ядра п р е дд в е р н о - ул итко- вого нерва (VIII, п. vestibulococh- learis, продолговатый мозг) — чувствительные. Улитковые ядра несут информацию от слуховых рецепторов, входят в состав слуховой сенсорной системы (п. 16.6). Вестибулярные ядра воспринимают информацию от рецепторов вестибулярного аппарата (вестибулярная система, см. п. 6.14 и 16.7). Ядра блуждающего нерва (X, п. vagus, продолговатый мозг). Заднее (парасимпатическое) ядро регулирует функции внутренних органов. Чувствительное ядро одиночного пути, иннервирующее слизистую оболочку нёба, корня языка, дыхательных путей, аортальное тельце, органы шеи, грудной и брюшной полости, участвует в качестве афферентного звена в глотательном, жевательном, дыхательных, висцеральных рефлексах. Двойное (двигательное) ядро обеспечивает сократительную активность мышц нёба, глотки, верхней части пищевода и гортани, реализует рефлексы глотания, рвоты, формирование голоса, кашля, чиханья. Ядра лицевого нерва (VII, л. facialis, мост). Двигательное ядро вызывает сокращения мимической и вспомогательной жевательной мускулатуры, регулирует передачу звуковых колебаний в среднем ухе в результате сокращения стремянной мышцы. Чувствительное ядро одиночного пути, иннервируя вкусовые луковицы передней 2/3 языка, участвует в анализе вкусовых раздражителей, в осуществлении моторных и секреторных пищеварительных рефлексов. Верхнее слюноотделительное (парасимпатическое) ядро стимулирует выделение секретов подъязычной, подчелюстной слюнных желез и слезной железы. Ядро отводящего нерва (VI, п. abducens, мост) обеспечивает сокращение наружной прямой мышцы глаза, в результате чего он поворачивается наружу. Ядра тройничного нерва (V, п. trigeminus) включают три чувствительных (среднемозговое, мостовое, спинномозговое) ядра и одно двигательное. Среднемозговое чувствительное ядро получает импульсацию от мышц языка, жевательных и мышц глазных яблок. Мостовое и спинальное чувствительные ядра получают тактильную, температурную, висцеральную и болевую импульсацию от первичных афферентных нейронов, расположенных в тройничном ганглии и иннервирующих кожу, слизистые оболочки, органы лица и головы. Информация, поступающая от них, обеспечивает формирование соответствующих ощущений и поведенческих реакций. Двигательное ядро тройничного нерва управляет главными жевательными мышцами; мышцами, напрягающими мягкое нёбо и барабанную перепонку; афферентную импульсацию оно получает от сенсорных ядер V нерва и нейронов РФ. Ядра глазодвигательного нерва (III, п. oculomotorius, средний мозг). Двигательное ядро обеспечивает сокращение верхней, нижней и внутренней прямой, нижней косой мышц глаза и мышцы, поднимающей верхнее веко. Добавочное (парасимпатическое) ядро, иннервируя ресничную мышцу и сфинктер зрачка, осуществляет соответственно рефлексы аккомодации глаза и сужения зрачка. Ядро блокового нерва (IV, п. trochlearis, средний мозг), иннервируя верхнюю косую мышцу, осуществляет поворот глаза кнаружи и вниз. Все центры ствола мозга целесообразно объединить в три группы. 1. Жизненно важные вегетативные центры регуляции систем: дыхания, сердечно-сосудистой системы, пищеварения. Поражение продолговатого мозга ведет к остановке дыхания и смерти. 2. Центры защитных рефлексов, в осуществлении которых участвуют по несколько черепных нервов, — это рефлексы чиханья, кашля, мигания, слезоотделения, рвоты. 3. Ядра, управляющие мускулатурой конечностей и туловища. Ядро шва (продолговатый мозг). Аксоны его нейронов поступают в спинной мозг, многие из них содержат серотонин. На интернейронах заднего рога этот путь образует тормозные синапсы (угнетение передачи сигналов), а на мотонейронах — возбуждающие. В целом, моноаминонергические пути модулируют реактивность нейронных цепей спинного мозга, а не конкретные движения. Вестибулярные ядра (мост) оказывают возбуждающее влияние на а- и у-мо- тонейроны спинного мозга мышц-раз- гибателей конечностей, туловища, шеи и тормозное влияние на мотонейроны сгибателей этих частей тела. Подобный эффект вызывают и ретикулярные ядра моста (медиальный ретикулоспинальный путь). Красные ядра (средний мозг, рубро- спинальный путь), напротив, оказывают тормозное влияние на мышцы-разгибатели и возбуждающее — на мышцы- сгибатели. Такое же влияние оказывает и ретикулярное гигантоклеточное ядро продолговатого мозга (латеральный ретикулоспинальный путь, рис. 6.16). Статические и статокинетические рефлексы, замыкающиеся на уровне
Мышца-разгибатель Рис. 6.16. Механизмы регуляции тонуса мышц и возникновения децеребрационной ригидности при перерезке ствола мозга между средним мозгом и мостом (пунктирная линия)'.
РФ — ретикулярная формация; Д — ядро Дейтерса; а- и у- — мотонейроны; вставочные нейроны спинного мозга: светлый — возбуждающий, черный — тормозной ствола мозга, регулируя тонус мышц, обеспечивают естественную позу, восстановление нарушенной позы и сохранение равновесия при перемещении организма в пространстве (Магнус Р., 1924). Статические рефлексы обеспечивают естественную позу в покое и восстановление нарушенной позы. Их разделяют на рефлексы позы (положения тела, с вестибулорецепторов) и выпрямительные рефлексы. Шейные рефлексы позы рассматривались в п. 6.12. Вестибулярные рефлексы позы направлены на сохранение естественной позы, они замыкаются на уровне моста. Для их выявления на шею животного накладывают гипсовую повязку, чтобы исключить рефлексы с проприорецеп- торов шеи. При естественном положении головы импульсация с рецепторов лабиринта минимальная. Если животное располагается спиной вниз, то тонус мышц-разгибателей становится максимальным (лабиринтный рефлекс); если животное располагается спиной вверх — минимальным. При вертикальной позе человека (голова вверх, шея несколько наклонена вперед) шейные и лабиринтные рефлексы усиливают друг друга, происходит повышение тонуса разгибателей нижних конечностей и сгибателей верхних конечностей. После выключения вестибулярного аппарата в эксперименте у животного вестибулярный рефлекс исчезает (опыт Р. Магнуса). Этот рефлекс сохраняется у бульбарного организма (перерезка ствола мозга между мостом и верхними бугорками четверохолмия среднего мозга), однако возникает резкое преобладание тонуса мышц-разгибателей и понижение тонуса мышц-сгибателей — децеребрационная ригидность (Шеррингтон Ч., 1896). Ригидность развивается вследствие преобладания возбуждающих нисходящих влияний на а- и у-мотонейроны над тормозными (см. рис. 6.16). Бульбарный организм не может поддерживать естественную позу. При наклоне головы влево или вправо повышается тонус мышц-разгибателей на соответствующей стороне (предотвращение падения вследствие смещения центра тяжести). При поражении лабиринта одного уха человек часто падает на эту сторону. Выпрямительные рефлексы наблюдаются у мезэнцефального организма при обязательном участии красного ядра (перерезка в эксперименте между средним и промежуточным мозгом). Благодаря этим рефлексам организм способен принимать естественную позу при ее нарушении, например, при положении животного на боку. Вначале выпрямляется голова (рефлексогенная зона — вестибулярный аппарат и кожа), затем — туловище (рефлексогенная зона — проприорецепторы мышц шеи и рецепторы кожи). При выключении этих рецепторов рефлексы исчезают. В условиях натуральной жизни важную роль в осуществлении выпрямительных рефлексов играет зрение. С т а т о к и н е т и ч е с к и е рефлексы возникают при ускорениях прямолинейного или вращательного движений организма. Сокращения мышц при этом направлены на преодоление действующих на человека ускорений, на сохранение нормальной позы, ориентации в пространстве. Они также осуществляются с помощью среднего мозга. Эти рефлексы запускаются с рецепторов вестибулярного аппарата. Они включают лифтные рефлексы, нистагм головы и глаз, перераспределение мышечного тонуса при ходьбе и беге. Лифтные рефлексы. При остановке движущегося вниз лифта и в начале быстрого подъема лифта, в нижних конечностях повышается тонус мышц-разгибателей, что обеспечивает преодоление сил инерции, хотя наблюдается некоторое сгибание конечностей под влиянием массы тела. В начале опускания лифта и при остановке движущегося вверх лифта наблюдаются противоположные явления. Правда, существует и другая точка зрения, учитывающая только внешние признаки — пассивное сгибание или разгибание конечностей. Глазной нистагм (пилообразные движения глаз), способствует сохранению зрительной ориентации. Нистагм имеет две фазы. При вращении сначала происходит медленное отклонение глазных яблок в сторону, противоположную вращению. Затем глаза быстро движутся обратно — по ходу вращения «догоняют организм». Медленное отклонение вызывается с рецепторов полукружных каналов, быстрый компонент связан с влиянием корковых центров (при наркозе он исчезает). Во врачебной практике направление нистагма обозначают по быстрому компоненту. Исследование нистагма (степени его выраженности, длительности) используется для оценки функционального состояния вестибулярного анализатора и дифференциальной диагностики поражения ЦНС. Перераспределение мышечного тонуса при ходьбе и беге обеспечивает сохранение равновесия и естественной позы. Например, при сгибании одной конечности тонус мышц-разгибателей другой конечности повышается. Мезэнцефалический организм имеет нормальный тонус мышц, но у него отсутствуют произвольные движения. Однако в среднем мозге, как и в спинном, имеются нейронные цепи, в которых запрограммированы ритмические движения конечностей (локомоция); они возникают при стимуляции соответствующих центров. В нормальных условиях их деятельность запускается корой большого мозга (см. п. 16.19, 6.21). 6.15. Проводниковая функция ствола мозга Как и в спинном мозге, в стволе мозга имеются восходящие и нисходящие пути. Восходящие пути ствола мозга подразделяют на специфические и неспецифические. Специфические пути включают медиальную и латеральную петли и другие пути. Медиальная петля формируется в основном аксонами нейронов клиновидного ядра (Бурдаха) и тонкого ядра (Голля); роль этих путей — см. табл. 6.2. Волокна медиальной петли переключаются преимущественно в вентральных задних специфических ядрах таламуса. В медиальную петлю входят также волокна от ядер V нерва (тригеминоталамический путь), проводящие импуль- сацию от головы (проприоцептивная и тактильная чувствительность). К медиальной петле на подходе к таламусу присоединяется часть волокон от ядра одиночного пути (вкусовая и висцеральная чувствительность) и небольшая часть волокон бокового спиноталамического пути (острая локализованная боль). Латеральная петля является составной частью слуховой системы, переключается в медиальном коленчатом теле. Пути зрительной системы переключаются в латеральном коленчатом теле. Часть афферентной импульсации поступает в кору мозжечка по дорсальному спинно-мозжечковому пути (Флекси- га) и переднему (вентральному) спинно-мозжечковому пути (Говерса, см. табл. 6.2), а также по вестибуломозжечковому пути от вестибулорецепторов. Из коры мозжечка обработанная импульса- ция передается по церебеллоталамиче- скому пути в двигательные вентральные ядра таламуса, откуда после переключения проецируется в соматосенсорную, моторную и премоторную зоны коры большого мозга. Неспецифичес к ие пути (экстралемнисковые) — эт
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2021-04-12; просмотров: 208; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 13.58.103.70 (0.022 с.) |