Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Координационная деятельность и интегративная роль центральной нервной системыСодержание книги
Поиск на нашем сайте
Координационная деятельность ЦНС — это согласование деятельности различных отделов ЦНС с помощью упорядочения распространения возбуждения между ними. Специальных центров координационной деятельности ЦНС не существует. Она осуществляется благодаря взаимодействию процессов возбуждения и торможения. Если выключить один из этих процессов, деятельность организма нарушается. Если выключить процесс торможения в ЦНС, например, введением стрихнина в опыте на лягушке (блокатора постсинаптического торможения), деятельность организма нарушается в результате беспрепятственной иррадиации по ЦНС процессов возбуждения. В этом случае нарушается двигательная активность из-за расстройства элементарной координации в деятельности нейронов ЦНС, ответственных за поочередное возбуждение и торможение спинальных мотонейронов, контролирующих работу мышц. При блокаде же процессов возбуждения в ЦНС, например, с помощью эфира лягушка становится обездвиженной, ее мышцы теряют тонус, активность лягушки полностью нарушается. Имеется ряд факторов, обеспечивающих взаимодействие центров между собой, центров и рабочих органов и формирующих приспособительную деятельность организма. Фактор структурно-функциональной связи — это наличие связи между отделами ЦНС, между ЦНС и различными органами, обеспечивающей преимущественное распро странение возбуждения между ними. Имеется несколько вариантов подобной связи. Прямая связь — это управление другим центром (ядром) или рабочим органом с помощью посылки к ним эфферентных импульсов (команд). Например, нейроны дыхательного центра продолговатого мозга посылают импульсы к а-мотонейронам спинного мозга, от которых нервные импульсы поступают к дыхательным мышцам, пирамидные клетки коры большого мозга посылают импульсы к нейронам спинного мозга. Обратная связь (обратная афферен- тация) — управление нервным центром или рабочим органом с помощью афферентных импульсов, поступающих от них. В данном случае центр имеет, естественно, и прямую связь с образованиями, функцию которых он контролирует, но обратная афферентация делает прямую связь более совершенной в функциональном отношении, (принцип обратной связи в регуляции функций организма). Если нарушить прямую связь центра с регулируемым центром или органом, то управление становится вообще невозможным. Если же нарушить только обратную связь, управление сильно страдает. Деафферентация конечности, например, ведет к нарушению управления ею. Если перерезать задние корешки спинного мозга, обеспечивающие чувствительную иннервацию одной из конечностей у собаки, то эта конечность может совершать движения в ритме дыхания и жевания. При нарушении обратной связи становится невозможной регуляция функций по отклонению (основной тип регуляции всех функций в организме). Денервация, например, аортальной и синокаротидной рефлексогенных зон (нарушение принципа обратной связи) ведет к развитию гипертонии — увеличению артериального давления.
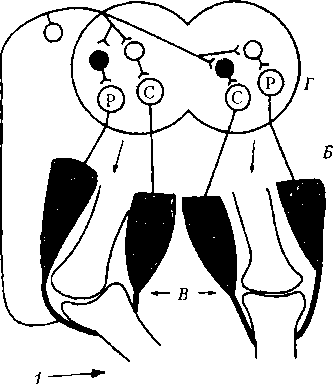
Возвратная связь — вид функциональной связи, обеспечивающей торможение нейронов вслед за их возбуждением. Ярким примером этого вида связи является торможение мотонейронов (иннервирующих мышцы конечностей) с помощью возвратного постсинаптического торможения. При этом вслед за возбуждением мотонейронов, обеспечивающих сокращение мышцы-сгибателя или мышцы-разгибателя, развивается торможение нейронов этого же центра, с помощью тормозных клеток Реншоу (см. рис. 6.8, А). Это обеспечивает расслабление мышцы и возможность очередного ее сокращения, например, при шагательном рефлексе и других повторяющихся сокращениях и расслаблениях мышц. Реципрокная (взаимная) связь — вид связи, обеспечивающей торможение центра-антагониста (рис. 6.11, А) при возбуждении центра-агониста. Например, при вызове сгибательного рефлекса конечности импульсы от рефлексогенной зоны (кожа) поступают через вставочные нейроны к мотонейронам центра мышц-сгибателей, а также одновременно — к центру-антагонисту (мышц-разгибателей), но с включением на пути тормозного нейрона, который образует тормозной синапс на нейронах центра-разгибателя. Мышцы-разгибатели поэтому не сокращаются и не препятствуют сгибанию конечности, что осуществляется с помощью прямого постсинаптического торможения (см. рис. 6.8, Г). Реципрокные взаимоотношения между центрами представлены довольно широко. Так, при возбуждении центра глотания тормозится центр жевания, возбуждение центра вдоха тор-
Рис. 6.11. Рефлексы нижних конечностей: А — дуга сгибательного (защитного) рефлекса; Б — дуга перекрестного разгибательного рефлекса; В — коленные суставы с мышцами; Г — сегмент спинного мозга; —> — раздражение кожи;? — афферентный путь; -I — эфферентные пути от а-мотонейронов центров сгибания (С) и разгибания (Р). Нейроны: светлые — возбуждающие, черные — тормозные
мозит центр выдоха, рефлекс глотания тормозит вдох. Принцип модульной (ансамблевой) структурно-функциональной организации ЦНС также следует рассматривать как один из факторов согласования деятельности различных отделов ЦНС. Каждый модуль (нейронный ансамбль) представляет собой совокупность повторяющихся локальных нейронных сетей, обрабатывающих и передающих информацию с помощью внутренних и внешних связей. Один модуль может входить в состав различных функциональных образований. Основным функциональным признаком модульной организации в деятельности мозга является локальный синергизм реакций нейронов центральной части ансамбля, окруженной зоной заторможенных нейронов, — тормозная окантовка (А. Б. Коган, О. Г. Чора- ян). Модули могут включать несколько нейронов, десятки, тысячи нейронов, при этом совокупность клеток, образующих модуль, обеспечивает появление у модуля новых свойств, которыми не обладают отдельные нейроны. Деятельность каждой клетки в составе модуля становится функцией не только поступающих к ней сигналов, но и функцией процессов, обусловленных той или иной конструкцией модуля (П.Г. Костюк). Фактор субординации также играет важную роль в координационной деятельности ЦНС — это подчинение нижележащих отделов ЦНС вышележащим. Например, ядро Дейтерса (продолговатый мозг), нейроны красного ядра (средний мозг) управляют активностью а- и у-мотонейронов спинного мозга. В процессе эволюции наблюдается тенденция к увеличению роли вышележащих отделов головного мозга в обеспечении координированной деятельности нижележащих центров (цефализация), причем с преобладанием тормозных влияний. Восходящие влияния преимущественно возбуждающие, что обеспечивает поддержание постоянной активности нервных центров, посылающих импульсы к нижележащим отделам ЦНС. Фактор силы процесса возбуждения также проявляется в согласовании деятельности нервных центров. При поступлении импульсов к одному и тому же центру от различных рефлексогенных зон {принцип общего конечного пути) центр реагирует на более сильное возбуждение. Например, слабое раздражение кожи туловища у собаки вызывает чесательный рефлекс нижней конечности — собака почесывает кожу. После прекращения действия слабого. раздражителя и окончания чесательного рефлекса наносят более сильное раздражение на эту же конечность, вызывающее оборонительный рефлекс (сгибание конечности), — организм избавляется от раздражителя. После окончания оборонительного рефлекса наносят одновременно два раздражения. В этом случае возникает только оборонительный рефлекс, чесательный рефлекс оказывается заторможенным. Таким образом, в борьбе за общий конечный путь побеждает более сильное возбуждение — выполняется более важная команда в биологическом отношении, что является важным приспособительным результатом.
Одним из факторов координационной деятельностиЦНСявляется одностороннее проведение возбуждения в химических синапсах ЦНС, так как это обеспечивает упорядоченное распространение возбуждения. В случае двустороннего проведения в синапсах наблюдалась бы иррадиация возбуждения в различных направлениях. Синаптическая потенциа- ция участвует в процессах обеспечения координационной деятельности ЦНС при выработке навыков, поскольку возбуждение распространяется в ЦНС быстрее и точнее по проторенным путям, возбудимость которых повышена (механизм — см. п. 6.5). Недостаточно координированные движения в начале выработки навыка постепенно становятся более точными — координированными. Дополнительные, ненужные движения постепенно устраняются. Доминанта также играет важную роль в координационной деятельности ЦНС, так как доминантное состояние двигательных центров обеспечивает автоматизированное выполнение двигательных актов, например, в процессе трудовой деятельности человека, при выполнении гимнастических элементов. При этом движения более точные, а также экономные в энергетическом и эмоциональном отношениях (подробнее об этом — см. п. 6.5). Интегративная роль нервной системы (от лат. integrum — цельный, объединение каких-либо частей) заключается в объединении органов и систем в единую функциональную систему — организм. Такое объединение становится возможным благодаря трем факторам: 1) участию ЦНС в управлении опорнодвигательным аппаратом с помощью соматической нервной системы; 2) регуляции функций всех тканей и внутренних органов с помощью ВНС и эндокринной системы; 3) наличию прямых и обратных (афферентных) связей ЦНС со всеми соматическими и вегетативными эффекторами. Можно выделить четыре основных уровня ЦНС, каждый из которых вносит свой вклад в обеспечение интегративной деятельности ЦНС. Первый уровень — нейрон. Взаимодействие возбуждающих и тормозящих входов, взаимодействие субсинаптических нейрохимических процессов в протоплазме в конечном итоге определяют, возникнет ли та или иная последовательность ПД на выходе нейрона или нет, т.е. будет ли дана команда другому нейрону, рабочему органу или нет, и возникнет ли соответствующая реакция. Таким образом, благодаря множеству возбуждающих и тормозящих синапсов на нейроне, он превратился в ходе эволюции в решающее устройство. Второй уровень — нейрональный ансамбль (модуль), обладающий качественно новыми свойствами, отсутствующими у отдельных нейронов — один и тот же модуль может участвовать в деятельности различных центров и отделов ЦНС и обеспечивать возникновение более сложных реакций ЦНС.
Третий уровень — нервный центр. Нервные центры часто выступают как автономные командные устройства, реализующие управление тем или иным процессом на периферии в составе саморегулирующейся, само- восстанавливающейся, самовоспроиз- водящейся системы, — организм. Это достигается за счет множественных прямых, обратных, реципрокных связей в ЦНС, наличия прямых и обратных связей с периферическими органами. Четвертый уровень — высший уровень интеграции, включающий все центры регуляции, объединяемые корой большого мозга в единую регулирующую систему,а отдельные органы и системы — в единую физиологическую систему — организм, подсистемы которого постоянно формируют функциональные системы. Это достигается взаимодействием главных систем ЦНС — лимбической формации, РФ, подкорковых образований и неокор- текса как высшего отдела ЦНС, организующего поведенческие реакции и их вегетативное обеспечение в различных условиях жизнедеятельности организма. 6.10. Структурнофункциональная характеристика спинного мозга Спинной мозг имеет сегментарное строение (31—33 сегмента), и каждый его сегмент связан с определенной частью тела. Спинной мозг включает пять отделов: шейный (С,-С8), грудной (Th,—Th|2), поясничный (L,—L5), крестцовый (S,— S5) и копчиковый (Со,—Со2). В процессе эволюции сформировалось два утолщения — шейное (сегменты, иннервирующие верхние конечности) и пояснично- крестцовое (сегменты, иннервирующие нижние конечности) — как результат повышенной нагрузки на эти отделы спинного мозга. В этих сегментах соматические нейроны наиболее крупные, их больше; в каждом корешке этих сегментов больше нервных волокон, они имеют наибольшую толщину. У некоторых видов животных подобные утолщения отсутствуют — например, у змеи, которая передвигается благодаря равномерному участию в процессе движения всей мускулатуры тела. Тренировка любого органа обеспечивает прогрессивное его развитие не только в фило-, но и в онтогенезе. Естественно, при этом совершенствуется и функция органа. Орган, который не получает достаточной нагрузки, постепенно атрофируется. В спинном мозге насчитывают около 13 млн нейронов (3 % — мотонейроны, 97 % — вставочные нейроны, в том числе относящиеся и к ВНС). При этом выделяют: 1) нейроны соматической и вегетативной нервной системы; 2) эфферентные, афферентные, вставочные, ассоциативные; 3) возбуждающие и тормозные. Афферентные нейроны соматической нервной системы локализуются в спинальных ганглиях и ганглиях черепных нервов. Окончания дендрита афферентного нейрона ветвятся на периферии и воспринимают изменения внешней или внутренней среды организма. Их отростки, проводящие афферентную импульсацию от мышечных, сухожильных и кожных рецепторов, вступают в ствол мозга и соответствующие сегменты спинного мозга и образуют синаптические контакты либо непосредственно на а-мотонейронах
(возбуждающие синапсы), либо на вставочных нейронах — возбуждающих и тормозных. Эфферентные нейроны спинного мозга, относящиеся к соматической нервной системе, включают а- и у-мотонейроны, являются эффекторными, поскольку иннервируют непосредственно рабочие органы — эффекторы (скелетные мышцы). а-мотонейроны иннервируют экстра- фузальные мышечные волокна (скелетные мышцы). Выделяют aj-мотонейро- ны — быстрые, иннервирующие белые мышечные волокна (лабильность — около 40 имп./с), и а2-мотонейроны — медленные, иннервирующие красные мышечные волокна (лабильность — 10— 15 имп./с). Низкая лабильность а-мото- нейронов объясняется длительной следовой гиперполяризацией, сопровождающей ПД. На одном а-мотонейроне насчитывается до 20 тыс. синапсов — от нисходящих путей вышележащих отделов ЦНС, кожных рецепторов, пропри- орецепторов. у-мотонейроны иннервируют ин- трафузальные мышечные волокна, т.е. волокна мышечных веретен (пропри- орецепторов). Эти нейроны обладают высокой лабильностью — до 200 имп./с, но их аксонаМ-Свойственна более низкая скорость проведения возбуждения — 15—40 м/с. у-мотонейроны локализуются между a-мотонейронами, участвуют в регуляции мышечного тонуса (см. п. 6.13 и 6.14). Вставочные (промежуточные) нейроны образуют связи с чувствительными нейронами, с мотонейронами ствола мозга и спинного мозга. Они обеспечивают связь спинного мозга с ядрами ствола мозга, а через них — с корой большого мозга. К ним также поступает импульсация от вышележащих отделов ЦНС. Этим нейронам присуща высокая лабильность —до 1 тыс. имп./с; они могут быть как возбуждающими, так и тормозными. Нейроны симпатического отдела ВНС расположены в боковых рогах грудного, поясничного и, частично, шейного отделов спинного мозга (С8—Ь2); являются также вставочными, фоновоактивны, частота их разрядов — 3-5 имп./с. Они регулируют функции всех внутренних органов, органов головы и сосудов. Нейроны парасимпатического отдела ВНС локализуются в сакральном отделе спинного мозга (S2—S4), они также вставочные, фоновоактивны. Их функция — регуляция деятельности нижнего отдела толстой кишки и мочеполовых органов. Центры спинного мозга участвуют в регуляции большинства внутренних органов и скелетных мышц; в частности, центр зрачкового рефлекса локализуется в сегментах С8—Th2, регуляции деятельности сердца — в сегментах Th]—Th5, слюноотделения — в сегментах Th2— Th5, регуляции функции желудка — в сегментах Th6—Th9, почек — в сегментах Th5—L3. Сегментарно расположены центры гладких мышц внутренних органов, центры пиломоторных рефлексов, регулирующие функции потовых желез и сосудов. Парасимпатическую иннервацию получают из спинного мозга (S2—S4) все органы малого таза: мочевой пузырь, часть толстой кишки ниже ее левого изгиба, половые органы. У женщин парасимпатическая иннервация обеспечивает сосудистые реакции клитора, влагалища; у мужчин — рефлекторный компонент эрекции. Центры управления скелетной мускулатурой находятся во всех отделах спинного мозга и иннервируют по сег- ментарному принципу мышцы шеи (С\- С4), диафрагмы (С3-С5), верхних конечностей (С5—Th2), туловища (Thj-Lj) и нижних конечностей (L2-S5). Повреждения определенных сегментов спинного мозга или его проводящих путей вызывают специфические расстройства чувствительности и двигательные нарушения. Каждый сегмент спинного мозга участвует в чувствительной иннервации трех дерматомов. Имеется дублирование и двигательной иннервации скелетных мышц, что повышает надежность функционирования опорно-двигательного аппарата и восприятия внешних раздражителей. РФ спинного мозга обнаруживается на уровне шейных сегментов между передними и задними рогами, на уровне верхнегрудных сегментов — между боковыми и задними рогами в белом веществе, примыкающем к серому. Ее нейроны имеют большое количество отростков. РФ участвует в регуляции тонуса мышц, вегетативных функций, афферентной импульсации. Спинной мозг имеет собственный ассоциативный аппарат, устанавливающий связь между сегментами и внутри сегментов. Ассоциативный аппарат спинного мозга участвует в регуляции движений конечностей и туловища, тонуса мышц, обеспечивая поддержание позы. Спинной мозг выполняет проводниковую и рефлекторную функции. 6.11. Проводниковая функция спинного мозга Общая характеристика. С помощью проводниковой функции спинного мозга вышележащие отделы ЦНС получают информацию от тела организма и внутренних органов, управляют скелетной мускулатурой тела и конечностей, регулируют функции внутренних органов (симпатическая нервная система — для всего организма, парасимпатическая — в основном для органов таза). Проводниковая функция спинного мозга осуществляется с помощью восходящих и нисходящих путей. Афферентная информация поступает в спинной мозг через задние корешки, эфферентная импульсация и регуляция функций различных органов и тканей организма осуществляются через передние корешки (закон Белла— Мажанди). Каждый корешок представляет собой множество нервных волокон. Например, дорсальный корешок кошки включает 12 тыс., а вентральный — 6 тыс. нервных волокон. Медиатором многих афферентных нейронов является глутамат, модуляторами — нейро- пептиды (вещество Р, ВИП, энкефалин). Все афферентные входы в спинной мозг несут информацию от трех групп рецепторов: 1) кожных рецепторов', болевых, температурных, прикосновения, давления, вибрации; щекотки; 2) проприорецепторов: мышечных (мышечных веретен), сухожильных (рецепторов Гольджи), надкостницы и оболочек суставов; 3) рецепторов внутренних органов — висцерорецепторов (механо-, осмо-, термо- и хеморецепторов). Значение афферентной импульсации, поступающей в спинной мозг: • несет информацию об изменениях окружающей среды; • участвует в координационной деятельности ЦНС по управлению скелетной мускулатурой: при выключении Таблица 6.2
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-04-12; просмотров: 305; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.137.175.166 (0.016 с.) |