Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
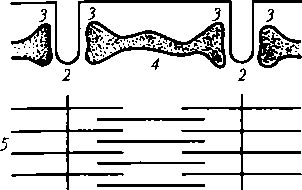
Назначение основных структурных элементов
Характеристика основных элементов мышечного волокна. От клеточной мембраны мышечного волокна (сарколеммы) вглубь отходят многочисленные поперечные инвагинации (Т-трубочки), которые обеспечивают ее взаимодействие с СПР (рис. 5.1). Последний представляет собой систему связанных друг с другом цистерн и отходящих от них в продольном направлении канальцев, расположенных между миофибриллами. Терминальные (концевые) цистерны СПР примыкают к Т-трубоч- кам, формируя так называемые триады. В цистернах содержится Са2+, играющий важную роль в мышечном сокращении. В саркоплазме имеются внутриклеточные элементы: ядра, митохондрии, белки (в том числе миоглобин), капельки жира, гранулы гликогена, фосфатсо- держащие вещества, различные малые молекулы и электролиты. Миофибриллы — субъединицы мышечного волокна. В одном мышечном волокне может насчитываться более 2 тыс. миофибрилл, их диаметр 1—2 мкм.
 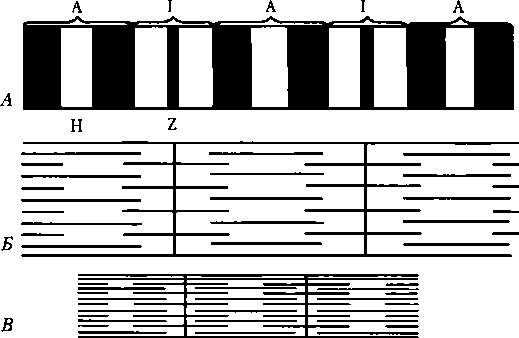
Элементы саркомера (см. рис. 5.3). Миозиновые протофибриллы образуют наиболее темную часть саркомера — A-диск (анизотропный, он сильно поляризует белый свет). Более светлый участок в центре A-диска называют Н-зоной. Светлый участок саркомера между двумя A-дисками называют 1-диском (изотропный, почти не поляризует свет). Он образован актиновыми протофибриллами, идущими в обе стороны от Z-полосок. Каждый саркомер имеет два набора тонких нитей, прикрепленных к Z-полоскам, и один комплект толстых нцтей, сосредоточенных в A-диске. В расслабленной мышце концы толстых и тонких филаментов в разной степени перекрывают друг друга на границе между А- и 1-дисками. Классификация мышечных волокон. По структурно-функциональным свойствам и цвету выделяют две основные группы мышечных волокон: быстрые и медленные. Белые (быстрые) мышечные волокна содержат больше миофибрилл и меньше — митохондрий, миоглобина и жиров, но больше гликогена и гликолитических ферментов; эти волокна называют гликолитическими. Капиллярная сеть, окружающая эти волокна, относительно редкая. Скорость рабочего цикла у данных волокон примерно в 4 раза больше, чем у медленных, что объясняется более высокой АТФазной активностью быстрых волокон, но они обладают малой выносливостью. У белых мышечных волокон число нитей актина и миозина больше, чем у красных, поэтому они толще и сила их сокращения больше, чем у красных волокон. Красные мышечные волокна содержат много митохондрий, миоглобина, жирных кислот. Эти волокна окружены густой сетью кровеносных капилляров, они имеют меньший диаметр. Митохондрии обеспечивают высокий уровень окислительного фосфорилирования, поэтому данные волокна называют оксидатпивными. Красные мышечные волокна подразделяются на две подгруппы: быстрые и медленные. Медленные волокна могут выполнять работу в течение относительно продолжительного периода времени; утомление в них развивается медленнее. Они более приспособлены к тоническим сокращениям. Красные быстрые волокна по скорости утомления занимают промежуточное положение между белыми и красными медленными. Скорость их сокращения близка к скорости сокращения белых волокон, что также объясняется высокой АТФазной активностью миозина красных быстрых волокон.
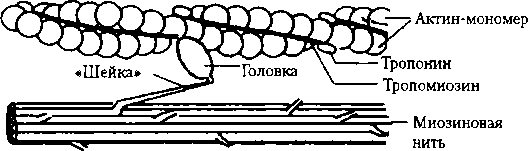
Также имеется незначительное число истинныхтонических мышечных волокон; на них локализуется по 7-10 синапсов, принадлежащих, как правило, нескольким мотонейронам, например, в глазодвигательных мышцах, мышцах среднего уха. ПКП этих мышечных волокон не вызывают генерации ПД в них, а непосредственно запускают мышечное сокращение. Группа мышечных волокон, иннервируемых ветвлениями аксона одного а-мотонейрона, формирует двигательную (нейромоторную) единицу. В мышцах, совершающих быстрые и точные движения, например в глазодвигательных, нейромоторные единицы состоят из 3—5 мышечных волокон. В мышцах, осуществляющих менее точные движения (например, мышцы туловища и конечностей), двигательные единицы включают сотни и тысячи мышечных волокон. Большая двигательная единица, по сравнению с малой, включает крупный мотонейрон с относительно толстым аксоном, который образует большое число концевых веточек в мышце и, следовательно, иннервирует большое число мышечных волокон. Все мышечные волокна одной двигательной единицы, независимо от их количества, относятся к одному типу. Все скелетные мышцы по своему составу являются смешанными, т.е. образованы красными и белыми мышечными волокнами. Специфическим свойством всех мышц является сократимость — способность сокращаться, т.е. укорачиваться или развивать напряжение. Реализация этой способности осуществляется с помощью возбуждения и его проведения по мышечному волокну (свойства соответственно возбудимости и проводимости). Скелетные мышцы не обладают ав- томатией, управляются организмом произвольно импульсацией из ЦНС, поэтому их называют также произвольными. Гладкие мышцы по собственному желанию не сокращаются, поэтому их называют также непроизвольными, но они обладают автоматией. Функции скелетной мышцы Обеспечение двигательной активности организма —поиск и добывание воды и пищи, ее захват, жевание, глотание, оборонительные реакции, трудовая деятельность — физическая и творческая работа художника, писателя, ученого, композитора в конечном итоге выражается в движении: рисование, письмо, игра на музыкальном инструменте и т.п. Обеспечение дыхания (движений грудной клетки и диафрагмы). Коммуникативная функция (устная и письменная речь, мимика и жесты). Участие в процессах терморегуляции организма с помощью изменения интенсивности сократительного термогенеза (п. 4.4). 5.2. Механизм сокращения скелетной мышцы Укорочение мышцы является результатом сокращения множества саркомеров. При укорочении актиновые нити скользят относительно миозиновых, в результате чего длина каждого саркомера мышечного волокна уменьшается. При этом длина самих нитей остается неизменной. Миозиновые нити имеют поперечные выступы (поперечные мостики) длиной около 20 нм. Каждый выступ состоит из головки, которая соеди-
Последовательность процессов, обеспечивающих сокращение мышечного волокна (электромеханическое сопряжение): 1. После генерации ПД в мышечном волокне вблизи синапса (за счет электрического полд ПКП) возбуждение распространяется по мембране миоцита, в том числе по мембранам поперечных Т-трубочек. Механизм проведения ПД по мышечному волокну такой же, как и по безмиелиновому нервному волокну — возникший ПД вблизи синапса посредством своего электрического поля обеспечивает возникновение новых ПД в соседнем участке волокна и т.д. (непрерывное проведение возбуждения). 2. Потенциал действия Т-трубочек за счет своего электрического поля активирует потенциалуправляемые кальциевые каналы на мембране СПР, вследствие чего Са2+ выходит из цистерн СПР согласно электрохимическому градиенту (см. рис. 5.1). Следует заметить, что существуют и другие точки зрения. Например, указывают на физическое или химическое сопряжение (Vander А., Sherman J., Luciano D., 2004) либо на механическое воздействие Т-трубочек на СПР. Однако, по нашему мнению, такие утверждения необоснованны, поскольку у Т-трубочек, представляющих собой впячивания клеточной мембраны внутрь клетки, при возникновении ПД размеры и форма не изменяются (мышечное волокно, как известно, сначала возбуждается, а потом сокращается). Сведений о выделении Т-тру- бочкой медиатора нет. Расстояние, на котором электрическое поле ПД может деполяризовать мембрану, до 1 мм (по аналогии с безмиелино- вым нервным волокном), что обеспечивает высокую надежность активации СПР.
3. В межфибриллярном пространстве Са2+ контактирует с тропонином, что приводит к его конформации и смещению тропомиозина, в результате чего на нитях актина обнажаются активные участки, с которыми соединяются головки миозиновых мостиков. 4. В результате взаимодействия с актином АТФазная активность головок миозиновых нитей усиливается, обеспечивая освобождение энергии АТФ, которая расходуется на сгибание миозинового мостика, внешне напоминающего движение весел при гребле (гребковое движение) (см. рис. 5.4), обеспечивающее скольжение актиновых нитей относительно миозиновых. На совершение одного гребкового движения расходуется энергия одной молекулы АТФ. При этом нити сократительных белков смещаются на 20 нм. Присоединение новой молекулы АТФ к другому участку головки миозина ведет к прекращению зацепления ее, но при этом энергия АТФ не расходуется. При отсутствии АТФ головки миозина не могут оторваться от актина — мышца напряжена; таков, в частности, механизм трупного окоченения. 5. После этого головки поперечных мостиков в силу своей эластичности возвращаются в исходное положение и устанавливают контакт со следующим участком актина; далее вновь происходит очередное гребковое движение и скольжение актиновых и миозиновых нитей. Подобные элементарные акты многократно повторяются. Одно гребковое движение (один шаг) вызывает уменьшение длины каждого саркомера на 1 %. При сокращении изолированной мышцы лягушки без нагрузки 50% укорочение саркомеров происходит за 0,1 с. Для этого необходимо совершение 50 гребковых движений. Миозиновые мостики сгибаются асинхронно, но в связи с тем, что их много и каждая миозиновая нить окружена несколькими актиновыми нитями, сокращение мышцы происходит плавно. Расслабление мышцы происходит благодаря процессам, протекающим в обратной последовательности. Ре поляризация сарколеммы и Т-трубочек ведет к закрытию кальциевых потенци- алуправляемых каналов мембраны СПР. Са-насосы возвращают Са2+ в СПР (активность насосов возрастает при увеличении концентрации свободных ионов). Снижение концентрации Са2+ в межфибриллярном пространстве вызывает обратную конформацию тропонина, в результате чего тропомиозиновые нити изолируют активные участки актиновых филаментов, что делает невозможным взаимодействие с ними головок поперечных мостиков миозина. Скольжение актиновых нитей вдоль миозиновых в обратном направлении происходит под действием сил гравитации и эластической тяги элементов мышечного волокна, что восстанавливает исходные размеры саркомеров. Источником энергии для обеспечения работы скелетных мышц является АТФ, расходы которой значительны. Даже в условиях основного обмена на функционирование мускулатуры организм затрагивает около 25 % всех своих энергоресурсов. Затраты энергии! резко возрастают во время выполнения физической работы.
Запасы АТФ в мышечном волокне незначительны (5 ммоль/л) и могут обеспечить не более 10 одиночных сокращений. Расход энергии АТФ необходим для осуществления следующих процессов. В о-пе р в ы х, энергия АТФ расходуется на обеспечение работы Na/K-насоса (он поддерживает градиент концентрации Na+ и К+ внутри и вне клетки, формирующих ПП и ПД, обеспечивающего электромеханическое сопряжение) и работы Са-насоса, который понижает концентрацию Са2+ в саркоплазме после сокращения мышечного волокна, что приводит к расслаблению. В о-вт о р ы х, энергия АТФ расходуется на гребковое движение миозиновых мостиков (сгибание их). Ресинтез АТФ осуществляется с помощью трех энергетических систем организма. 1. Фосфогенная энергетическая система обеспечивает ресинтез АТФ за счет имеющегося в мышцах высокоэнергоемкого КФ и образовавшейся при расщеплении АТФ аденозиндифосфор- ной кислоты (аденозиндифосфат, АДФ) с образованием креатина (К): АДФ + + КФ АТФ + К. Это мгновенный ресинтез АТФ, при этом мышца может развивать большую мощность, но кратковременно — до 6 с, поскольку запасы КФ в мышце ограниченны. 2. Анаэробная гликолитическая энергетическая система обеспечивает ресинтез АТФ за счет энергии анаэробного расщепления глюкозы до молочной кислоты. Этот путь ресинтеза АТФ является быстрым, но тоже кратковременным (1—2 мин), так как накопление молочной кислоты тормозит активность гликолитических ферментов. Однако лактат, вызывая местный сосудорасширяющий эффект, улучшает кровоток в работающей мышце и снабжение ее кислородом и питательными веществами. 3. Аэробная энергетическая система обеспечивает ресинтез АТФ с помощью окислительного фосфорилирования углеводов и жирных кислот, протекающего в митохондриях мышечных клеток. Этот способ может обеспечить энергией работу мышц в щечение нескольких часов и является основным способом энергетического обеспечения работы скелетных мышц. 5.3. Виды мышечных сокращений В зависимости от характера сокращений мышцы различают три их вида: изометрическое, изотоническое и аук- сотоническое. Ауксотоническое сокращение мышцы заключается в одновременном изменении длины и напряжения мышцы. Этот вид сокращения характерно для натуральных двигательных актов и бывает двух видов: эксцентрическое, когда напряжение мышцы сопровождается ее удлинением — например, в процессе приседания (опускания), и концентрическое, когда напряжение мышцы сопровождается ее укорочением — например, при разгибании нижних конечностей после приседания (подъем). Изометрическое сокращение мышцы — когда напряжение мышцы возрастает, а длина ее не изменяется. Этот вид сокращения можно наблюдать в эксперименте, когда оба конца мышцы зафиксированы и отсутствует возможность их сближения, и в естественных условиях — например, в процессе приседания и фиксации положения. Изотоническое сокращение мышцы заключается в укорочении мышцы при ее постоянном напряжении. Этот вид сокращения возникает, когда сокращается ненагруженная мышца с одним закрепленным сухожилием, не поднимая (не перемещая) никакого внешнего груза либо поднимая груз без ускорения. В зависимости от длительности сокращений мышцы выделяют два их вида: одиночное и тетаническое. Одиночное сокращение мышцы возникает при однократном раздражении нерва или самой мышцы. Обычно мышца укорачивается на 5—10 % от исходной длины. На кривой одиночного сокращения выделяют три основных периода: 1) латентный — время от момента нанесения раздражения до начала сокращения; 2) период укорочения (или развития напряжения)', 3) период расслабления. Продолжительность одиночных сокращений мышц человека ва-
Рис. 5.5. Соотношение времени возникновения ПД (Л) и одиночного сокращения (Б) медленного волокна скелетной мышцы теплокровного. Стрелка — момент нанесения раздражения. Время сокращения быстрых волокон в несколько раз короче
риабельна. Например, у камбаловидной мышцы она составляет 0,1 с. В латентный период возникает возбуждение мышечных волокон и его проведение вдоль мембраны. Соотношения длительности одиночного сокращения мышечного волокна, его возбуждения и фазовые изменения возбудимости мышечного волокна показаны на рис. 5.5. Длительность сокращения мышечного волокна значительно дольше таковой ПД потому, что необходимо время на работу Са-насосов для возвращения Са2+ в СПР и окружающую среду и большей инерционности механических процессов по сравнению с электрофизиологическими. Тетаническое сокращение — это длительное сокращение мышцы, возникающее под действием ритмического раздражения, когда каждое последующее раздражение или нервные импульсы поступают к мышце, пока она еще не расслабилась. В основе тетанического сокращения лежит явление суммации одиночных мышечных сокращений (рис. 5.6) — увеличение амплитуды и длительности сокращения
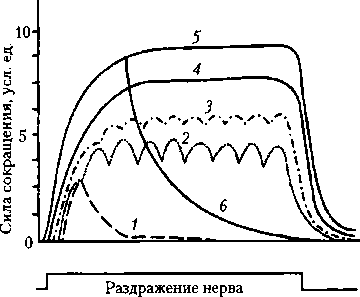
Рис. 5.6. Суммация сокращений икроножной мышцы лягушки:
1 — кривая одиночного мышечного сокращения в ответ на первое раздражение расслабленной мышцы; 2 — кривая одиночного сокращения той же мышцы в ответ на второе раздражение; 3 — кривая суммированного сокращения, полученного в результате спаренного раздражения сокращающейся мышцы (обозначено стрелками) при нанесении на мышечное волокно или целую мышцу двух и более быстро следующих друг за другом раздражений. При этом раздражения должны поступать в период предыдущего сокращения. Увеличение амплитуды сокращений объясняется увеличением концентрации Са2+ в гиалоплазме при повторном возбуждении мышечных волокон, поскольку Са-помпа не успевает возвращать его в СПР. Са2+ обеспечивает увеличение числа зон зацепления миозиновых мостиков с нитями актина. Если повторные импульсы или раздражения поступают в фазу расслабления мышц, возникает зубчатый тетанус. Если повторные раздражения приходятся на фазу укорочения, возникает гладкий тетанус (рис. 5.7). Амплитуда сокращения и величина напряжения, развиваемые мышечными волокнами при гладком тетанусе, обычно в 2—4 раза больше, чем при одиночном сокращении. Тетаническое сокращение мышечных волокон, в отличие от одиночных сокращений, быстрее вызывает их утомление. При возрастании частоты стимуляции нерва или мышцы амплитуда гладкого тетануса увеличивается. Максимальный
Рис. 5.7. Сокращения икроножной мышцы лягушки при различной частоте раздражения седалищного нерва:
1 — одиночное сокращение (частота 1 Гц); 2, 3 — зубчатый тетанус (15-20 Гц); 4, 5 — гладкий тетанус (25-60 Гц); 6 — расслабление при песси- мальной частоте раздражения (120 Гц) тетанус получил название оптимума. Увеличение тетануса объясняется накоплением Са2+ в гиалоплазме. При дальнейшем увеличении частоты стимуляции нерва (около 100 Гц) мышца расслабляется вследствие развития блока проведения возбуждения в нервно-мышечных синапсах — пессимум Введенского (частота раздражения пессимальная) (см. рис. 5.7). Пессимум Введенского можно получить и при прямом, но более частом раздражении мышцы (около 200 имп./с), однако при этом для чистоты эксперимента следует заблокировать нервно-мышечные синапсы. Если после возникновения пессимума уменьшить частоту стимуляции до оптимальной, то амплитуда мышечного' сокращения мгновенно возрастает — свидетельство того, что пессимум не является результатом утомления мышцы или истощением энергетических ресурсов. В естественных условиях отдельные мышечные волокна чаще сокращаются в режиме зубчатого тетануса, однако сокращение целой мышцы напоминает гладкий тетанус, вследствие асинхронности их сокращения. 5.4. Показатели физической деятельности мышц Сила мышцы Единицы измерения. В системе СИ сила выражается в ньютонах (Н). В физиологической практике силу мышцы, как правило, определяют по максимальной массе груза, который может быть поднят при ее сокращении. В условиях целостного организма определяют «становую», «кистевую» силу, силу сгибателей и т.п. Факторы, определяющие силу мышцы. Анатомическое строение: перистые мышцы (волокна расположены косо, под углом к продольной оси) способны развивать гораздо большее напряжение, чем мышцы с параллельным расположением волокон. В связи с этим принято определять так называемое физиологическое поперечное сечение мышцы, т.е. сумму поперечных сечений всех волокон, из которых состоит мышца. У перистых мышц физиологическое поперечное сечение значительно превосходит анатомическое (геометрическое). К числу наиболее сильных относятся жевательные мышцы. Выделяют понятие «удельная сила мышцы» — отношение общей силы мышцы в ньютонах к физиологическому поперечному сечению мышцы (Н/см2). Удельная сила находится в пределах 50— 150 Н/см2. Удельную силу мышцы выражают также и в килограммах на квадратный сантиметр (кг/см2). Так, для трехглавой мышцы она составляет 17 кг/см2, для сгибателя плеча — 8 кг/см2, для икроножной мышцы — 1 кг/см2, для гладкой мышцы — 1 кг/см2. В разных мышцах тела соотношение между числом медленных и быстрых мышечных волокон неодинаково и очень сильно отличается у разных людей, а также в разные периоды жизни. Одиночное мышечное волокно способно развивать напряжение 0,1—0,2 Н. Исходная длина мышцы тоже влияет на силу ее сокращения. При умеренном предварительном растяжении мышцы сила ее сокращения увеличивается, а при сильном растяжении она уменьшается, вплоть до отсутствия сокращения из-за отсутствия зон зацепления между нитями актина и миозина. При оптимальной длине (в состоянии покоя), при которой все головки миозиновых нитей способны контактировать с актиновыми филаментами, сила мышечного сокращения вырастает максимально. Предварительное растяжение мышцы увеличивает ее эластическую тягу, что также ведет к увеличению последующего ее сокращения. Это осуществляется за счет белка титина, нити которого одним концом прикреплены к Z-пластинке, другим — к миозину и растягиваются подобно пружине. При сильном укорочении мышцы уменьшается (по непонятным причинам) сродство тропонина к Са2+, что ограничивает максимальную силу сокращений. Число возбужденных волокон также влияет на силу одиночного сокращения мышцы. Оно определяется силой раздражения в эксперименте или числом возбужденных мотонейронов в натуральных условиях. Сила тетанического сокращения мышцы зависит от степени выраженности суммации сокращений в каждом мышечном волокне, что определяется частотой импульсации — она возрастает до оптимума. Работа мышцы (А). В механике работа определяется как произведение силы (/), приложенной к телу, на расстояние (Z) его перемещения под воздействием данной силы: А = /’х L (Дж). Утомление мышцы. При мышечной работе у человека со временем развивается утомление — сила мышечных сокращений постепенно уменьшается, и в конечном итоге наступает момент, когда человек уже не в состоянии продолжать работу. Скорость развития утомления зависит от ритма работы и величины груза. Большой груз или слишком частый ритм работы приводят к быстрому развитию утомления, в результате чего выполненная работа бывает ничтожна. Наибольшей бывает работа при некотором среднем, оптимальном для данного человека, ритме работы и среднем, оптимальном грузе (правило средних нагрузок). При любой силе изометрического сокращения мышцы работа равна нулю, несмотря на расход энергии и развивающееся утомление. Причиной утомления является накопление К+ в Т-трубочках (при частых сокращениях), накопление молочной кислоты, расход энергетического материала. Мощность мышцы (работа, совершаемая в единицу времени) в системе СИ выражается в ваттах (Дж/с2). Максимальная мощность соответствует выполнению наибольшего объема работы в течение минимального отрезка времени. Однако в этом случае быстро развивается утомление. 5.5. Структурнофункциональные особенности гладких мышц Расположение актина и миозина в гладких мышцах не столь упорядочено, Z-мем- браны и саркомеры в них отсутствуют, поэтому при микроскопическом исследовании не выявляется характерная для скелетной мышцы поперечная исчерчен- ность, что и определяет название этих мышц — гладкие. Форма гладкомышечных клеток веретенообразная, диаметр волокна в утолщенной части составляет 2-10 мкм, длина 50-400 мкм. В клетке имеется одно ядро, митохондрий относительно мало. СПР представлен плоскими везикулами, расположенными в непосредственной близости от внутренней поверхности клеточной мембраны. Он содержит мало ионов Са2+. Нервно-мышечные синапсы отличаются от таковых у исчерченных мышц, причем наиболее ярко отличие выражено у симпатической нервной системы. Постганглионарные волокна (аксона ганглионарных симпатических нейронов) по своему ходу среди миоцитов образуют многочисленные утолщения (расширения), из которых выделяется медиатор. Последний диффундирует в межклеточном пространстве и взаимодействует с постсинаптическими рецепторами, которые располагаются равномерно по всей мембране гладкомышечных клеток, что ведет к стимуляции или угнетению функций органа (например, торможение моторики кишки, усиление работы сердца, сужение кровеносного сосуда). В гладких мышцах бронхов и крупных артерий нервное влияние передается без генерации ПД, сокращение этих мышц обеспечивают ВПСП.
|
|||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2021-04-12; просмотров: 181; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.219.22.169 (0.039 с.) |