Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Особенности свойств гладких мышцСодержание книги
Поиск на нашем сайте
Возбудимость. Потенциал покоя большинства гладкомышечных клеток составляет —60—70 мВ, у миоцитов, обладающих спонтанной активностью, — —30—60 мВ. Потенциал действия более продолжителен (10—50 мс), чем у скелетных мышц — до 10 мс. У некоторых миоцитов после начальной быстрой реполяризации формируется плато, которое удлиняет ПД до 500 мс; оно связано с поступлением в клетку Na+ и Са2+. Деполяризация мембраны обусловлена в основном диффузией Са 2+ в клетку. Проводимость. Структурнофункциональной единицей гладких мышц является пучок мышечных волокон. Взаимодействие между отдельными миоцитами осуществляется благодаря щелевым контактам, обладающим низким электрическим сопротивлением, и близко расположенным контактирующим элементам соседних мышечных волокон. Благодаря этому электрическое поле одной клетки в пучке обеспечивает возбуждение другой. Поэтому изолированно отдельные гладкомышечные клетки пучка не возбуждаются. Скорость распространения ПД в пределах пучка составляет 5-10 см/с. Причем для возбуждения всех миоцитов пучка не достаточно возбуждения одного миоцита (необходимо первоначальное возбуждение нескольких клеток). Сократимость. Сокращения гладкой мышцы определяются описанным выше характером распространения возбуждения — пучок гладкомышечных волокон сокращается как единое целое (пучок — функциональная единица гладкой мышцы). Активность гладкомышечной АТФазы миозина в 40—80 раз ниже активности АТФазы миозина исчерченной мышцы. Чем больше АТФаз- ная активность миозина, тем быстрее сокращается мышечное волокно. Поэтому гладкая мышца сокращается гораздо медленнее, чем скелетная. По этой же причине на сокращение гладкой мышцы меньше расходуется АТФ (экономичность). Кроме того, гладкая мышца не утомляется во время продолжительной активности — она приспособлена к длительному поддержанию тонуса. Главной особенностью электромеханического сопряжения в гладкой мышце является то, что основную роль в сопряжении играет входящий в клетку (при ее возбуждении) Са2+, поскольку его запасы в СПР гладкомышечных миоцитов незначительны. Другая важная особенность заключается в том, что регуляторным белком гладкой мышцы является кальмодулин (наличие тропонина не установлено), который связывается с Са2+. Комплекс Са2+ — кальмодулин активирует особый фермент (киназу легких цепей миозина), который переносит фосфатную-группу с АТФ на головку поперечного мостика миозина. Фосфорилированная головка миозина взаимодействует с актином. Это ведет к конфирмационным изменениям миозиновых мостиков, что обеспечивает скольжение нитей актина относительно нитей миозина.
Сокращение гладких мышц может быть результатом и химиомеханическо- го сопряжения (без формирования ПД), вследствие взаимодействия медиатора с мембранными рецепторами и активации различных ферментных систем, вызывающих взаимодействие актина и миозина, что и обеспечивает сокращение мышцы. Расслабление гладкомышечных миоцитов обусловлено инактивацией кальциевых каналов вследствие восстановления исходных значений МП. Активация кальциевого насоса в мембране миоцита и СПР обеспечивает выведение Са2+ в СПР и из гиалоплазмы клетки и снижение его концентрации, в результате чего инактивируется киназа легких цепей миозина, что приводит к прекращению фосфорилирования миозиновых головок, а следовательно, они утрачивают способность взаимодействовать с актином. Автоматия присуща клеткам — водителям ритма (пейсмекерам). В ее основе лежит спонтанно возникающая медленная деполяризация (препотенциал) — при достижении КП возникает ПД. Спонтанная деполяризация преимущественно обусловлена диффузией Са2+ в клетку. Частота генерируемых ПД зависит от скорости медленной деполяризации и соотношения МП и КП: чем меньше МП, тем ближе он к КП, и при этом легче возникают ПД. Автоматия практически не выражена у гладких мышц артерий, семенных протоков, радужки, ресничных мышц. Их функции полностью определяются ВНС. Пластичность выражается в том, что при растяжении гладких мышц их напряжение первоначально увеличивается, а затем снижается до исходного уровня. Таким образом, свойство пластичности проявляется в том, что гладкая мышца может не изменять напряжения как в укороченном, так и в растянутом состоянии. Эта особенность гладкой мышцы предотвращает избыточный рост давления в полых внутренних органах при их наполнении (мочевой пузырь, желудок и др.).
Однако растяжение гладкой мышцы может вызывать активацию процессов сокращения. Этот феномен, в частности, характерен для артериол, что является одним из важных механизмов регуляции их тонуса и регионарного кровотока в некоторых органах (мозг, почки, сердце). Стимуляция сокращения в этом случае происходит в результате того, что при растяжении пейсмекерных клеток активируются механоуправляемые каналы, в результате чего возникает ПД, который посредством своего электрического поля и щелевых контактов обеспечивает возникновение ПД в соседних клетках. Чрезмерное растяжение мочевого пузыря также вызывает его сокращение и эвакуацию мочи. Подобная реакция наблюдается при денервации органа и фармакологической блокаде внутриорганной системы. 97 Энергетическое обеспечение сокращения гладких мышц также осуществляется за счет молекул АТФ, ресинтез которой происходит, в основном, посредством анаэробного гликолиза.
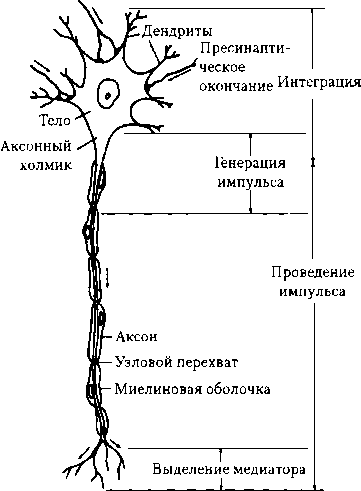
6.1. Структурнофункциональная характеристика центральной нервной системы Нервная клетка (нейрон) является структурной и функциональной единицей ЦНС. Она состоит из сомы (тела клетки с ядром) и отростков, представляющих собой большое число дендритов и один аксон (рис. 6.1). Любая деятельность самой ЦНС реализуется в конечном итоге с помощью функционирования отдельных клеток. ЦНС человека содержит свыше 100 млрд нервных клеток, взаимодействие между которыми осуществляется посредством множества синапсов. Число последних в тысячи раз больше количества самих клеток (1015—1016), так как их аксоны многократно делятся дихотомически. Нейроны оказывают влияние на органы и ткани также посредством синапсов. Важную роль в деятельности нервной системы играют глиальные клетки и ликвор. Развитие неврологических нарушений у больных СПИДом связано с гибелью 40—50 % корковых нейронов. Нервные клетки имеются и вне ЦНС: нейроны ганглиев ВНС, афферентные нейроны спинномозговых ганглиев и ганглиев черепных нервов. Однако периферических нервных клеток, по сравнению с центральными, мало — всего лишь около 25 млн. В теле нейрона синтезируются макромолекулы, которые транспортируются по аксону и дендритам; оно выполняет трофическую функцию по отношению к своим отросткам и клеткам-эффекторам. Отросток, лишенный связи с сомой нейрона, дегенерирует. ПП нейрона составляет —60—80 мВ, ПД — 80—110 мВ. Скопления тел нейронов образуют серое вещество мозга, отростков — белое вещество. Сома и дендриты каждого нейрона покрыты нервными окончаниями — синаптическими бутонами других нервных клеток и отростками глиальных клеток. На одном мотонейроне число синаптических бутонов может достигать 10—20 тыс. (см. рис. 6.1). Аксон начинается от тела клетки аксонным холмиком. Диаметр тела клетки составляет 10—100 мкм, аксона — 1—6 мкм; на периферии длина аксона может достигать 1 м и более. По аксону передаются нервные импульсы к другим нейронам и эффекторным клеткам.
Рис. 6.1. Структурно-функциональные элементы мотонейрона спинного мозга (по Р. Эккерт, Д. Рэнделл, Дж. Огастин, 1991, с изменениями)
Нейрон имеет большое число дендритов, на которых локализуется большинство синапсов нейрона, обеспечивающих (вместе с синапсами сомы) восприятие информации от других нервных клеток.
Основные функции нейрона — восприятие, переработка и хранение информации, передача сигнала другим нервным клеткам, регуляция деятельности клеток различных органов и тканей организма. Отдельные структурные элементы нейрона выполняют определенные функции. Структурами, воспринимающими импульсы от других нервных клеток, являются тело и дендриты нейрона с расположенными на них шипиками. Если Шипики не получают импульсацию, то они исчезают. Импульсы могут поступать и к окончанию аксона — аксо- аксонные синапсы, например в случае пресинаптического торможения. Число дендритов у нейрона весьма вариабельно — от 1 до 1500. Структура, где начинает формироваться ПД, — аксонный холмик. Структура, проводящая возбуждение к другому нейрону или к эффекторной клетке, — аксон. Структура, передающая импульсы на другую клетку, — это пресинаптиче- ский аппарат, представляющий собой нервное окончание с пресинаптиче- ской мембраной. В нем синтезируется и запасается медиатор. При возбуждении нервного окончания медиатор выбрасывается в синаптическую щель через пресинаптическую мембрану, которая обратно захватывает неиспользованный медиатор в нервное окончание. В процессе деятельности нервной системы отдельные нейроны образуют нейронные цепи (например, рефлекторная дуга), объединяются в ансамбли (модули), нейронные сети. Последние могут включать несколько нейронов, десятки, тысячи нейронов, при этом совокупность клеток, образующих модуль, обеспечивает появление новых свойств, которыми не обладают отдельные нейроны. Деятельность каждой клетки в составе модуля становится функцией не только поступающих к ней сигналов, но и функцией процессов, обусловленных той или иной конструкцией модуля (П. Г. Костюк); Примером восходящих сетей являются сенсорные афферентные пути. Глиальные клетки (нейроглия — «нервный клей»; от греч. neuron — нерв, glia — клей) более многочисленны, чем нейроны, составляют около 50 % объема ЦНС; способны к делению в течение всей жизни. По размерам глиальные клетки в 3—4 раза меньше нервных. Число их огромно — 14—1010 (в 10 раз больше, чем нейронов), с возрастом оно увеличивается (число нейронов уменьшается). Глиальные клетки окружают тела нейронов и их аксоны. Мембранный потенциал клеток нейроглии составляет -70—90 мВ. Электрические поля их ЛП взаимодействуют с электрическими полями ВПСП и ПД нейронов. Глиальные клетки выполняют н е- сколько функций: изолирующую (глиоциты окутывают нейроциты в виде своеобразного футляра и продуцируют миелин), опорную, защитную (выработка цитокинов, фагоцитоз, при повреждении нейронов образуют глиальный рубец), обменную (синтез белков памяти, снабжение нейронов питательными веществами, поддержание оптимальной концентрации ионов К+ в интерстиции). Микроглиальные клетки способны к ритмическому изменению своего объема (период «сокращения»— 1,5 мин, «расслабления» — 4 мин). Циклы изменения объема повторяются через каждые 2—20 ч. Полагают, что пульсация способствует продвижению аксоплазмы в нейронах и влияет на ток межклеточной жидкости. Норадреналин усиливает, серотонин угнетает пульсацию глиоцитов. Глиоциты, охватывая снаружи капилляры ЦНС, участвуют в формировании ГЭБ.
Ликвор — бесцветная прозрачная жидкость, заполняющая мозговые желудочки, спинномозговой канал и субарахноидальное пространство. Происхождение ликвора связано с интерстициальной жидкостью мозга, значительная часть его (ликвора) образуется сосудистыми сплетениями желудочков мозга — это совокупность фильтрата плазмы крови и интерстициальной жидкости. От плазмы крови ликвор отличается, как и межклеточная жидкость других тканей, низким содержанием белка (0,1 г/л, а в плазме крови 75 г/л), меньшим содержанием аминокислот (0,8 и 2,0 ммоль/л соответственно) и глюкозы (3,9 и около 5,0 ммоль/л соответственно). Его объем 100-200 мл (12—14 % общего объема мозга), за сутки вырабатывается около 600 мл. Обновление этой жидкости происходит 4-8 раз в сутки; давление ликвора при горизонтальном положении человека составляет 7—14 мм рт. ст., при вертикальном — 15—30 мм рт. ст. Функции ликвора: выполняет защитную роль (она обладает бактерицидными свойствами, содержит! иммуноглобулины G и А, систему ком-/ племента, моноциты и лимфоциты, является своеобразной гидравлической «подушкой» мозга). Интерстициальная жидкость является непосредственной питательной средой клеток мозга, в нее клетки выделяют также и продукты своего обмена. Отток ликвора выполняет дренажную функцию (препятствие оттоку ликвора в кровь может сопровождаться тяжелыми нарушениями в деятельности ЦНС). Отток ликвора осуществляется несколькими путями: 30—40 % — через субарахноидальное пространство в продольный синус венозной системы головного мозга; 10—20 % — через периневральные пространства черепных и спинномозговых нервов в лимфатическую систему; часть ликвора реабсорбируется сосудистыми сплетениями мозга. Нейроны ЦНС классифицируют по нескольким признакам. По влиянию на другие клетки имеются возбуждающие и тормозные нейроны. По виду м ед и ат о р а, выделяющегося в окончания аксонов, различают нейроны серотонинергические, адренергические, холинергические и др. По специфичности воспринимаемой сенсорной информации нейроны высших отделов ЦНС могут быть моно-, би- и полисенсорными. Например, моносенсорными являются нейроны первичных сенсорных зон в коре большого мозга. Первичные афферентные нейроны (спинальных ганглиев и ганглиев черепных нервов) также моносенсорны: они воспринимают с помощью собственных рецепторов только один вид раздражителя (адекватного) — свет, химическое вещество, электрическое, механическое и температурное воздействие. Бисенсорные нейроны встречаются во вторичных сенсорных зонах в коре большого мозга. Полисенсорные нейроны — это нейроны ассоциативных зон мозга (третичные сенсорные зоны), моторной коры, они реагируют на любые раздражители (звук, свет, прикосновение и др.).
По активности имеются фоновоактивные и «молчащие» нейроны, возбуждающиеся только в ответ на раздражение. Фоновоактивные нейроны играют важную роль в поддержании тонуса ЦНС и, особенно, коры большого мозга. Фоновоактивные нейроны различаются по общему рисунку генерации импульсов, так как одни нейроны разряжаются непрерывно (ритмично или аритмично), другие — «пачками» импульсов. По направлению передачи информации нейроны подразделяют на: 1) афферентные, воспринимающие с помощью рецепторов информацию о внешней и внутренней среде организма и передающие ее в вышележащие отделы ЦНС; 2) эфферентные, передающие информацию к рабочим органам — эффекторам; нервные клетки, иннервирующие эффекторы, иногда называют эффекторными (эффекторные нейроны спинного мозга — мотонейроны делят на а- и у-мотонейроны); 3) вставочные (интернейроны), обеспечивающие взаимодействие между нейронами ЦНС различных уровней. В зависимости от отдела нервной системы выделяют нейроны соматической и вегетативной нервной системы. Синапсы ЦНС классифицируют также по нескольким признакам. По эффекту, как и нейроны, — возбуждающие и тормозные. В зависимости от местоположения — аксосоматические, аксодендритные, аксо-аксонные, дендро-дендритные, дендросоматические. По механизму передачи сигнала к другим клеткам: 1) химические синапсы (наиболее распространенный способ); 2) электрические синапсы (редко); 3) смешанные синапсы (электрохимические), которые обнаружены между нейронами латерального вестибулярного ядра; 4) часть нейронов гипоталамуса информацию передают с помощью гормонов. Химические синапсы имеют относительно широкую синаптическую щель (20-50 нм). В пресинаптической терминали содержится большое число пузырьков — пресинаптических везикул (диаметр около 50 нм), заполненных медиатором — химическим посредником. Медиатор либо образуется в теле нейрона и транспортируется в пресинаптиче- ское окончание аксонным транспортом, либо синтезируется непосредственно в пресинаптическом окончании и накапливается в везикулах. Для синтеза медиатора нужны ферменты, образующиеся в теле клетки на рибосомах. Медиатор в пресинаптическом окончании находится в двух пулах— готовом к экзоцитозу (пул 1, медиатор лежит на пресинаптической мембране изнутри) и мобилизационном (пул 2, находится на цитоскелете), из которого пополняется в течение нескольких секунд пул 1. Этот процесс активируется ионами Са2+. Химические синапсы передают сигнал относительно медленно, односторонне, менее надежно, чем электрические синапсы. Химическим синапсам присуще явление суммации. Электросекреторное сопряжение в синапсах ЦНС, как и в нервно-мышечных синапсах, обеспечивается ионами Са2+ — при возбуждении пресинаптической терминали открываются потенциалуправляемые Са-каналы, дополнительно Са2+ выделяется из эндоплазматического ретикулума. Кальций входит в нервное окончание согласно электрохимическому градиенту и запускает процесс высвобождения медиатора в синаптическую щель с помощью экзоцитоза — ион Са2+ активирует белковый экзоцитозный аппарат пресинапса, представляющий собой совокупность белков (синапсин, спектрин), активация которых обеспечивает выход медиатора в синаптическую щель. Синаптическая щель химических синапсов содержит поперечно расположенные гликопротеидные филаменты, обеспечивающие связи пре- и постсинаптической мембран и направление диффундирующего медиатора от пресинаптической мембраны к постсинаптической. Выделившийся в синаптическую щель медиатор взаимодействует с белком-рецептором постсинаптической мембраны, что и приводит нервную клетку в состояние возбуждения или торможения. Электрические синапсы имеются возбуждающие и тормозные (последних мало, изучены они недостаточно). Электрические синапсы имеют щель на порядок меньшую (2-4 нм), чем у химических синапсов, проводят сигнал в обе стороны без синаптической задержки, передача не блокируется при удалении Са2+; они малочувствительны к фармакологическим препаратам и ядам, практически не утомляемы, как и нервное волокно. Контактирующие мембраны клеток в составе электрического синапса связаны друг с другом полуканалами белковой природы — коннексонами (англ, connection — связь). Очень низкое удельное сопротивление сближенных пре- и постсинаптических мембран обеспечивает хорошую электрическую, проводимость — за счет электрического поля. Электрические возбуждающие синапсы обнаружены в головном мозге млекопитающих в составе мезэнцефального ядра тройничного нерва, вестибулярного ядра Дейтерса, ядра нижней оливы продолговатого мозга. Имеются следующие электрические синапсы: аксон—сома; аксон—дендрит; аксон—аксон; дендрит—дендрит; дендрит—сома; сома—сома. Электрические синапсы характерны для сердца, гладких мышц, секреторных клеток. Через коннексоны нейроны ЦНС обмениваются некоторыми компонентами цитоплазмы — пептидами, РНК, метаболитами, циклическими нуклеотидами, аминокислотами. Электрические синапсы оказывают влияние на метаболизм контактирующих клеток. Работа электрических синапсов может регулироваться близлежащими химическими синапсами. Например, между шипиками клеток ядра нижней оливы продолговатого мозга передача возбуждения блокируется, если выделяется медиатор в соседнем химическом синапсе. По механизму активации постсинаптические рецепторы химических синапсов делятся на два класса: 1) ионотропные, активация ионных каналов которых осуществляется нейромедиатором непосредственно с помощью G-белка (например, N-холино- рецептор); 2) метаботропные (например, а(- и а2-адренорецепторы), когда ионные каналы активируются через G-белок с помощью вторых посредников (цАМФ, цГМФ, ИФ3, ДАГ). После каждого цикла передачи сигнала выделившийся медиатор инактивируется специфическим ферментом (например, ацетилхолин — ацетилхолинэстеразой, норадреналин — моноаминоксидазой (МАО)), обратно захватывается пресинапсом, часть медиатора рассеивается. Функции ЦНС. Нервная система выполняет интегрирующую роль. Она объединяет в единое целое все ткани, органы организма, координируя их специфическую активность в составе целостных гомеостатических и поведенческих функциональных систем (П. К. Анохин). Основные частные функции нервной системы Обеспечение, сознания и всех видов психической деятельности. Управление деятельностью опорно-двигательного аппарата. Нервная система регулирует тонус мышц и посредством его перераспределения поддерживает естественную позу, а при нарушении последней восстанавливает ее; инициирует все виды движений в процессе трудовой деятельности, занятий физкультурой и спортом. Регуляция работы внутренних органов, осуществляемая с помощью ВНС и эндокринных желез. При этом в покое обеспечивается гомеостазис (постоянство внутренней среды организма), а во время работы — приспособительное регулирование деятельности внутренних органов согласно потребностям организма и поддержание гомеостазиса. Формирование целенаправленного поведения организма в социальном обществе и окружающей среде. Оно реализуется с помощью реакций избегания или избавления от неприятных раздражителей (защитные реакции организма), регуляции интенсивности обмена веществ при изменении температуры окружающей среды. Изменения внутренней среды организма, воспринимаемые субъективно в виде ощущений (жажда, голод), также побуждают организм к той или иной целенаправленной деятельности. 6.2. Медиаторы и рецепторы центральной нервной системы Медиатор (лат. mediator — посредник) — химическое вещество, с помощью которого сигнал передается от одной клетки к другой. В головном мозге к настоящему времени обнаружено около 30 БАВ. Вещество, из которого образуется медиатор (предшественник медиатора), попадает в сому или аксон из крови и ликвора, в результате биохимических реакций под действием ферментов превращается в соответствующий медиатор, затем транспортируется в синаптические везикулы. Медиатор может синтезироваться в теле нейрона или его окончании. При передаче сигнала с нервного окончания на другую клетку медиатор высвобождается в синаптическую щель и действует на рецептор постсинаптической мембраны. Как отмечалось выше, по механизму реагирования на медиатор все эффекторные рецепторы подразделяют на ионотропные и метаботропные. Большинство ионотропных и метаботропных рецепторов связано с G-белками (ГТФ-свя- зывающие белки). При действии медиатора на ионотропные рецепторы открываются ионные каналы непосредственно с помощью G-белка, и вследствие движения ионов в клетку или из клетки формируются ВПСП или ТПСП. Ионотропные рецепторы называют также рецепторами быстрого ответа (например, N-холино- рецептор, ГАМК^-, глицино-, 5-HT3(S3)- серотонинорецепторы). При действии медиатора на метаботропные рецепторы ионные каналы активируются через G-белок с помощью вторых посредников. Далее формируются ВПСП, ПД, ТПСП (электрофизиологические явления), с помощью которых запускаются биохимические (метаболические) процессы; при этом возбудимость нейрона и амплитуда ВПСП могут быть повышенными в течение секунд, минут, часов и даже дней. Вторые посредники могут также изменять активность ионных каналов.
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-04-12; просмотров: 111; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.12.34.211 (0.014 с.) |