Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Структурно-функциональная схема зрительного анализатора. Кодирование информации в воспринимающей, проводящей и центральной частях анализатора.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте Билет 1 1. Место физиологии в высшем медицинском образовании. Предмет и методы физиологии. Физиология — наука о природе, о существе жизненных процессов. Физиология изучает жизнедеятельность организма и отдельных его частей: клеток, тканей, органов, систем. Предметом изучения физиологии являются функции живого организма, их связь между собой, регуляция и приспособление к внешней среде, происхождение и становление в процессе эволюции и индивидуального развития особи. Физиологическая функция — проявления жизнедеятельности организма и его частей, имеющие приспособительное значение и направленные на достижение полезного результата. В основе функции лежит обмен веществ, энергии и информации. Физиология - это экспериментальная наука. Она использует два основных метода: наблюдение и эксперимент. Она изучает функции и процессы, протекающие в организме, отдельных органах и системах органов, механизмы их формирования, реализации и регуляции, изучает процессы – т.е. динамику явлений, состояний во времени и пространстве. Физиология относится к разряду фундаментальных наук. А это значит, что физиология изучает законы жизнедеятельности. Это значит, что она изучает наиболее важные взаимосвязи в живой материи. Она является базой для целой группы биологических прикладных наук, а именно - патологической физиологии, фармакологии. Физиологию определяют как теоретическую основу медицины: - Во-первых, это обусловлено тем, что физиология изучает процессы нормальной жизнедеятельности. Предметом интереса медицины являются болезни- больной человек и болезни. Чтобы понять отклонение, надо понимать нормальное течение процессов. - Во-вторых, физиология дает нормы для медицины, т.е. параметры нормальной деятельности органов и систем органов. - В-третьих, физиология дает методы оценки функций, т.е. она дает медицине практические методы диагностики. Основная задача физиологии — дальнейшее раскрытие специфических физиологических законов деятельности нервной системы и других систем организма человека и животных, что необходимо для разработки способов управления всеми проявлениями жизни организма, и прежде всего обмена веществ и энергии, психики и поведения. Следовательно, физиология участвует в выяснении сущности явлений жизни, изучения физики и химии живого, разработке способов управления жизненными процессами, в частности обменом веществ, наследственностью и направленными изменениями организмов. Цели физиологии как науки: - исследование законов осуществления нормальных функций в живом организме в зависимости от постоянно изменяющихся и развивающихся условий его жизни - исследование исторического, филогенетического, и индивидуального, онтогенетического, развития функций живого организма и их взаимосвязи. Для нормальной жизнедеятельности необходимо выполнение трех условий: - 1-ое условие нормальной жизнедеятельности - постоянство внутренней среды. - 2-ое - постоянный обмен внутренней среды организма веществом, энергией и информацией со средой окружения, внешней средой. Из биофизики известно - человеческий организм - открытая система. Следовательно, обмен идет в обе стороны: и туда и обратно. - 3-ий принцип жизнедеятельности, нарушение которого несовместимо с жизнью - адекватное приспособление организма к изменяющимся условиям внешней среды или среды обитания.
Структурно-функциональная схема зрительного анализатора. Кодирование информации в воспринимающей, проводящей и центральной частях анализатора. Зрительный анализатор - сложная нервно-рецепторная система человека, осуществляющая восприятие и анализ зрительных раздражений. Зрительный анализатор включает в себя три отдела: периферический (глаз), проводниковый (зрительный нерв, зрительные пути и подкорковые нервные образования) и центральный (зрительная зона коры головного мозга)
Билет 2 Билет 3 Механизмы цветовосприятия. Цветовое зрение, цветовосприятие, способность глаза человека и многих видов животных с дневной активностью различать цвета, т. е. ощущать отличия в спектральном составе видимых излучений и в окраске предметов. В глазу человека содержатся два типа светочувствительных клеток (рецепторов): высокочувствительные палочки, отвечающие за сумеречное (ночное) зрение, и менее чувствительные колбочки, отвечающие за цветное зрение. В сетчатке глаза человека есть три вида колбочек, максимум чувствительности которых приходится на красный, зелёный и синий участок спектра, то есть соответствует трем «основным» цветам. Они обеспечивают распознавание тысяч цветов и оттенков. Кривые спектральной чувствительности трёх видов колбочек частично перекрываются. Очень сильный свет возбуждает все 3 типа рецепторов, и потому воспринимается, как излучение слепяще-белого цвета (эффект метамерии). Равномерное раздражение всех трёх элементов, соответствующее средневзвешенному дневному свету, также вызывает ощущение белого цвета. В основе цветового восприятия лежит свойство света вызывать определенное зрительное ощущение в соответствии со спектральным составом отражаемого или испускаемого излучения. Цвета разделяют на хроматические и ахроматические. Хроматические цвета обладают тремя основными качествами: цветовым тоном, который зависит от длины волны светового излучения; насыщенностью, зависящей от доли основного цветового тона и примесей других цветовых тонов; яркостью цвета, т.е. степенью близости его к белому цвету. Различное сочетание этих качеств дает большое разнообразие оттенков хроматического цвета. Ахроматические цвета (белый, серый, черный) различаются лишь яркостью. При смешении двух спектральных цветов с разной длиной волны образуется результирующий цвет. Каждый из спектральных цветов имеет дополнительный цвет, при смешении с которым образуется ахроматический цвет белый или серый. Многообразие цветовых тонов и оттенков может быть получено оптическим смешением всего трех основных цветов красного, зеленого и синего. Количество цветов и их оттенков, воспринимаемых глазом человека, необычайно велико и составляет несколько тысяч.
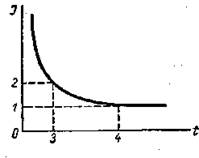
Билет 4 1. Возбудимость, количественная мера возбудимости. Кривая "сила-длительность". Реобаза. Хронаксия. Минимальный градиент. Лабильность. В озбудимость - способность отвечать на раздражение формированием электрической активности (потенциалом действия). У различных тканей возбудимость различна. У одной ткани возбудимость может изменяться в процессе жизнедеятельности, возбудимость у живой возбудимой ткани есть всегда, в не зависимости от действия раздражителя. Возбуждение - это состояние, это реализованная возбудимость. Количественной мерой возбудимости является порог раздражения — минимальная величина раздражителя, способная вызвать ответную реакцию ткани. Возбудимость проявляется в процессах возбуждения, которые представляют собой изменение процессов обмена веществ в клетках нервной ткани. Реобаза - наименьшая сила постоянного электрического тока, вызывающая при достаточной длительности его действия возбуждение в живых тканях. Понятие реобазы и хронаксии ввёл в физиологию Л. Лапик в 1909, определяя зависимость между силой тока и длительностью его действия при изучении наименьшего (порогового) эффекта возбудимых тканей. Реобаза, как и хронаксия, даёт представление о возбудимости тканей и органов по порогу силы и длительности действия раздражения. Реобаза соответствует порогу раздражения и выражается в вольтах или миллиамперах. Значение реобазы можно вычислить по формуле: i = a/t + b, где i - сила тока, t - длительность его действия, а и b - константы, определяемые свойствами ткани. Константа b является Р., так как при длительном действии раздражающего тока отношение a/t будет очень мало и i практически равняется b. Хронаксия - наименьшее время действия на ткань постоянного электрического тока удвоенной пороговой силы (удвоенной реобазы), вызывающего возбуждение ткани. Было также экспериментально установлено, что величина стимула, вызывающего возбуждающий эффект в тканях, находится в обратной зависимости от длительности его действия и графически выражается гиперболой - кривая <сила - время. Кривая «сила — длительность» по Лапику, Вейссу и Гоорвегу: 0—1 — реобаза, 0—3 — хронаксия. 0—4.—полезное время; по оси абсцисс — длительность раздражения, по оси ординат — сила раздражения (в условных единицах)
Лабильность (Н. Е. Введенский)- свойство возбудимой ткани воспроизводить максимальное число потенциалов действия в единицу времени. Максимальная лабильность - у нервной ткани. Частота раздражений, вызывающая максимальную реакцию называется оптимальной, а вызывающая угнетение реакции - пессимальной. Минимальный градиент – это минимальная скорость нарастания раздражителя, при которой ткань еще способна ответить возбуждением на данный раздражитель
Билет 5 1. Реакции невозбудимых и возбудимых мембран на раздражители, градуальность и закон "всё или ничего". Под действием многих раздражителей изменяется уровень мембранного потенциала. Такая реакция невозбудимых мембран связана, как правило, с изменением их ионной проницаемости за счет открывания или закрывания неспецифических потенциалнезависимых каналов. Между сдвигом мембранного потенциала невозбудимой мембраны и интенсивностью раздражителя, вызвавшего его, существует пропорциональная зависимость. Это свойство получило название градуалъности. Оно присуще невозбудимой мембране во всем диапазоне изменений разности потенциалов на ней. Градуальность свойственна и возбудимой мембране, но только в том случае, если трансмембранная разность потенциалов, изменяясь под действием раздражителя, не достигает критического мембранного потенциала. Такие раздражители, не доводящие деполяризацию до критического мембранного потенциала, и, следовательно, не вызывающие потенциала действия, называются подпороговыми для возбудимой ткани. Стимул, вызывающий сдвиг мембранного потенциала до КМП, считается пороговым, поскольку под действием его возникает потенциал действия (возбуждение). Раздражитель может иметь разную природу (механическую, химическую, электрическую и т. д.), но пороговым он будет тогда, когда сдвинет уровень мембранного потенциала от потенциала покоя до критического мембранного потенциала: Un = |ПП| - |КМП|. Надпороговые (более сильные) раздражители возбудимой мембраны тем более вызывают потенциал действия. Понятно, что градуальность характерна и для отклонений трансмембранной разности потенциалов (от уровня потенциала покоя) в сторону, противоположную деполяризации (в аксоне кальмара от -85 до -90 мВ и более). Такой сдвиг мембранного потенциала называют гиперполяризацией. Следовательно, градуальность присуща возбудимой мембране при любой ее гиперполяризации и при подпороговой (до критического мембранного потенциала) деполяризации. Иллюстрацией сказанному служит реакция возбудимой мембраны на пропускание через нее слабого постоянного электрического тока. Под катодом развивается деполяризация, а под анодом — гиперполяризация. Де- и гиперполяризационные сдвиги мембранного потенциала равны по абсолютной величине. Они тем больше, чем сильнее пропускаемый ток. Однако градуальность деполяризационных ответов под катодом свойственна только подпороговым электрическим стимулам, не приводящим к возбуждению. Не вызывая возбуждения, подпороговый электрический ток изменяет возбудимость возбудимой мембраны. Изменения возбудимости под действием подпорогового электрического тока называются электротоническими явлениями. Их установил в 1859 г. Э. Пфлюгер. Как уже говорилось, под действием порогового и надпорогового раздражителей возбудимая мембрана генерирует потенциал действия. Для этого процесса характерен закон ≪все или ничего≫. Он является антитезой градуальности. Смысл закона состоит в том, что параметры потенциала действия (амплитуда, длительность, фронты) не зависят от интенсивности раздражителя. Как только достигается критический мембранный потенциал, изменения разности потенциалов на возбудимой мембране определяются только свойствами ее потенциалзависимых ионных каналов, которые обеспечивают входящий ток (из межклеточной среды в цитоплазму). Среди них внешний стимул открывает только самые чувствительные. Другие открываются за счет предыдущих, уже независимо от раздражителя. Говорят о спонтанном (самопроизвольном) характере процесса вовлечения в трансмембранный перенос ионов все новых потенциалзависимых ионных калалов. Поэтому амплитуда, длительность, крутизна переднего и заднего фронтов потенциала действия зависят только от ионных градиентов на клеточной мембране и кинетических характеристик ее каналов. Закон ≪все или ничего≫ — характернейшее свойство одиночных клеток и волокон, обладающих возбудимой мембраной. Большинству многоклеточных образований он не свойствен, поскольку разные клетки даже в одном органе имеют неодинаковый порог возбуждения. Чем больше клеток вовлекают в возбуждение, тем сильнее ответ многоклеточной структуры. Исключение составляют структуры, организованные по типу синцития. Их возбуждение подчиняется закону ≪все или ничего≫.
Билет 6 Рефрактерность. Рефрактерность - временное снижение возбудимости ткани, возникающее при появлении потенциала действия. В этот момент повторные раздражения не вызывают ответной реакции (абсолютная рефрактерность). Она длится не более 0,4 миллисекунды, а затем наступает фаза относительной рефракторности, когда раздражение может вызвать слабую реакцию. Эта фаза сменяется фазой повышенной возбудимости - супернормальности. Показатель рефрактерности (рефрактерный период) - время, в течение которого возбудимость ткани снижена. Рефрактерный период тем короче, чем выше возбудимость ткани. Процесс возбуждения сопровождается изменением возбудимости. Таков смысл свойства рефрактерности. Это слово, в переводе означающее невпечатлительность, ввел в науку Э. Ж. Марей, обнаруживший в 1876 г. угнетение возбудимости миокарда в момент его возбуждения. Позднее рефрактерность была выявлена во всех возбудимых тканях. В1908 г. Н. Е. Введенский установил, что вслед за угнетением наступает некоторое повышение возбудимости возбужденной ткани. Выделяют три основные стадии рефрактерности, их принято называть фазами: - Развитие возбуждения вначале сопровождается полной утратой возбудимости (е = 0). Это состояние называют абсолютно рефрактерной фазой. Она соответствует времени деполяризации возбудимой мембраны. В течение абсолютно рефрактерной фазы возбудимая мембрана не может генерировать новый потенциал действия, даже если на нее подействовать сколь угодно сильным раздражителем (S„-> оо). Природа абсолютно рефрактерной фазы состоит в том, что во время деполяризации все потенциалзависимые ионные каналы находятся в открытом состоянии, и дополнительные стимулы не могут вызвать воротный процесс (им просто не на что действовать). - О тносительно рефрактерная фаза – возвращает возбудимостьот нуля к исходному уровню (е0). Относительно рефрактерная фаза совпадает с реполяризацией возбудимой мембраны. С течением времени во все большем числе потенциалзависимых ионных каналов завершаются воротные процессы, с которыми было связано предшествующее возбуждение, и каналы вновь обретают способность к следующему переходу из закрытого в открытое состояние под действием очередного стимула. Во время относительно рефрактерной фазы пороги возбуждения постепенно снижаются (S„o <S„ <со) и, следовательно, возбудимость восстанавливается до исходного уровня (0 < е < е0). - Ф аза экзальтации, для которой характерна повышенная возбудимость (е > е0). Она, очевидно, связана с изменением свойств сенсора напряжения во время возбуждения. За счет перестройки конформации белковых молекул изменяются их дипольные моменты, что приводит к повышению чувствительности сенсора напряжения к сдвигам мембранного потенциала (критический мембранный потенциал приближается к потенциалу покоя). Разным возбудимым мембранам присуща неодинаковая продолжительность каждой фазы рефрактерности. Так, в скелетных мышцах АРФ длится в среднем 2,5 мс, ОРФ — около 12 мс, ФЭ — приблизительно 2 мс. Миокард отличается гораздо более продолжительной АРФ — 250—300 мс, что обеспечивает четкую ритмичность сердечных сокращений и является необходимым условием жизни. В типичных кардиомиоцитах относительно рефрактерная фаза длится около 50 мс, а в сумме продолжительность абсолютно рефрактерной и относительно рефрактерной фаз примерно равна длительности потенциала действия. Различия в длительности рефракторных фаз обусловлены неодинаковой инерционностью потенциалзависимых ионных каналов. В тех мембранах, где возбуждение обеспечивается натриевыми каналами, рефрактерные фазы наиболее быстротечны и потенциал действия наименее продолжителен (порядка единиц миллисекунд). Если же за возбуждение ответственны кальциевые каналы (например, в гладких мышцах), то рефрактерные фазы затягиваются до секунд. В сарколемме кардиомиоцитов присутствуют и те, и другие каналы, вследствие чего длительность рефрактерных фаз занимает промежуточное значение (сотни миллисекунд).
Билет 7 Билет 8 Билет 9 Билет 10 Билет 11 Билет 12 Билет 13 1. Нервные волокна, их классификация, законы проведения возбуждения по нервным волокнам. Нервные волокна представляют собой отростки нейронов, которые покрыты глиальными оболочками. Оболочки нервных волокон в различных отделах нервной системы существенно разнятся по своему строению, что делит все волокна на безмиелиновые и миелиновые. Но все они состоят из отростка нервной клетки, находящегося в центре волокна, и в связи с этим имеет название осевого цилиндра (аксон), а также из глиальной оболочки, окружающей его. - Нервные волокна типа А делятся на четыре подгруппы: α, β, γ и δ. Они покрыты миелиновой оболочкой. Наиболее толстые из них — так называваемые альфа-волокна (Аα) — у теплокровных животных и человека имеют диаметр 12—22 мк и характеризуются значительной скоростью проведения возбуждения: 70—120 м/сек. Такие волокна проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам («двигательные» волокна) и от рецепторов мышц к соответствующим нервным центрам. Потенциал действия волокон Аα у теплокровных длится 0,5 мсек. После его окончания развивается следовой отрицательный потенциал, который продолжается 15—20 мсек и переходит в следовой положительный потенциал длительностью около 40—60 мсек. - Три другие группы волокон типа А: бета (Aβ)-, гамма (Аγ)- и дельта волокна имеют меньший диаметр — от 2 до 12 мк, меньшую скорость проведения и более длительный потенциал действия. Это преимущественно чувствительные волокна, проводящие возбуждение от различных рецепторов (тактильных, некоторых болевых, температурных и рецепторов внутренних органов) в центральную нервную систему. Исключение составляют лишь гамма-волокна, значительная часть которых проводит возбуждение в центробежном направлении — от клеток спинного мозга к так называемым интрафузальным мышечным волокнам которых расположены чувствительные нервные окончания — мышечные веретена. -- К волокнам типа В относятся миелиновые, преимущественно преганглионарные волокна автономной нервной системы. Скорость проведения возбуждения в этих волокнах у теплокровных животных составляет 3—14 м/сек. Продолжительность потенциала действия волокон типа В примерно в 2 раза превышает длительность потенциала действия волокон типа А. В среднем она составляет 1,2 мсек. Отличительной особенностью этих волокон является то, что в них не обнаруживается следового отрицательного потенциала: фаза реполяризации непосредственно переходит в следовой положительный потенциал, имеющий большую длительность — 100—300 мсек. - К волокнам типа С относят безмякотиые нервные волокна очень малого диаметра (порядка 0,5 мк). Скорость проведения возбуждения в этих волокнах 2—6 м/сек. Большинство С-волокон относится к постганглионарным волокнам симпатической нервной системы. К С-волокнам относят также те нервные волокна, которые участвуют в проведении возбуждения от болевых рецепторов и некоторых рецепторов холода, тепла и давления. Проведение возбуждения по нервам подчиняется следующим законам: - Закон анатомической и физиологической целостности нерва. Первая нарушается при перерезке, вторая – действии веществ, блокирующих проведение, например новокаина. - Закон двустороннего проведения возбуждения. Оно распространяется в обе стороны от места раздражения. В организме чаще всего возбуждение по афферентным путям оно идет к нейрону, а по эфферентным – от нейрона. Такое распространение называется ортодромным. Очень редко возникает обратное, или антидромное, распространение возбуждения. - Закон изолированного проведения. Возбуждение не передается с одного нервного волокна на другое, входящее в состав этого же нервного ствола. - Закон бездекрементного проведения. Возбуждение проводится по нервам без декремента, т.е. затухания. Следовательно, нервные импульсы не ослабляются проходя по ним. Скорость проведения прямо пропорциональна диаметру нерва.
Билет 14 Билет 15 Билет 16 Билет 17 1. Иррадиация и концентрация возбуждения в ЦНС. Механизмы взаимодействия нервных центров (принципы координации рефлекторной деятельности: реципрокность, общий конечный путь, субординация, обратная афферентация, доминанта) Иррадиация возбуждения. При сильном и длительном раздражении импульсы, поступившие в нервную систему, не ограничиваются возбуждением одного рефлекторного центра, а распространяются на другие центры. Чем сильнее раздражение, тем больше нервных центров охватывает иррадиация. Возбуждение большого количества различных нервных центров позволяет отобрать из них наиболее нужные и установить между ними новые функциональные связи — условные рефлексы. Большинство двигательных условных рефлексов формируется благодаря иррадиации возбуждения. Излишняя иррадиация возбуждения центральной нервной системы нарушает уравновешенность поведенческих реакций, приводит к расстройству рефлекторной деятельности. Иррадиация возбуждения ограничивается и уравновешивается торможением. Основными принципами координации являются: - Принцип сопряженного торможения или реципрокности - базируется на описанном выше одноименном виде торможения и означает возникновение при возбуждении одного центра одновременного торможения другого центра для обеспечения цепных рефлексов и сложной функции. Примером реципрокности может быть регуляция спинным мозгом противоположных по функциональному назначению мышц конечностей. Так, при возбуждении мотонейронов, иннервирующих мышцы сгибатели правой ноги, реципрокно тормозятся мотонейроны мышц разгибателей этой ноги и возбуждаются мотонейроны мышц разгибателей левой ноги. Формирующийся цепной характер рефлексов вслед за этим вызывает возбуждение мотонейронов разгибателей правой ноги и реципрокно — торможение мотонейронов сгибателей правой ноги и возбуждение мотонейронои сгибателей левой ноги. Таким образом, реципрокные взаимоотношения между указанными рефлексами обеспечивают цепной шагательный рефлекс. Реципрокные взаимоотношения имеют место и между рефлексами вдоха и выдоха, когда возбуждение центра вдоха тормозит центр выдоха и наоборот, что обеспечивает ритмичную смену фаз в процессе внешнего дыхания. - Принцип доминанты - был открыт А.А.Ухтомским. Доминантой называют обший принцип деятельности нервной системы, проявляющийся в виде господствующей в течение определенного времени системы рефлексов, реализуемых доминирующими центрами, которые подчиняют себе или подавляют деятельность других нервных центров и рефлексов. Нейроны доминирующих центров приобретают более низкий уровень критической деполяризации мембран, т.е. становятся более возбудимыми, и способны эффективнее осуществлять пространственную и временную суммацию нервных импульсов. Синаптическое проведение к этим нейронам облегчено и поэтому они могут возбуждаться и за счет «посторонних» импульсов от не имеющих прямых связей с доминирующими центрами информационных каналов. Вследствие суммации многочисленных ВПСП возбуждение нейронов как и число возбужденных клеток в доминирующем центре нарастает и осуществляемые им рефлекторные реакции легко реализуются. Преобладание рефлексов доминирующего центра над другими рефлекторными актами становится особенно выраженным, поскольку через систему вставочных нейронов доминирующий центр сопряженно тормозит другие центры и текущие рефлексы. Принцип доминанты позволяет концентрировать внимание и строить поведение для достижения определенной намеченной цели. - Принцип общего конечного пути - описанный Ч.Шеррингтоном, рассматривался при изложении процесса конвергенции возбуждения. - Принцип субординации нервных центров - (принцип подчинения) проявляется в виде регулирующего влияния выше расположенных нервных центров на ниже расположенные. Так, двигательные центры головного мозга управляют спинальными мотонейронами. Примером такого влияния является феномен центрального торможения спинальных рефлексов открытый И.М.Сеченовым и получивший название сеченовского торможения. В эксперименте И.М.Сеченова раздражение зрительных бугров лягушки с помощью кристаллика поваренной соли (т.е. раздражение ретикулярной формации среднего мозга) приводило к торможению спинальных двигательных рефлексов, вызываемых погружением лапки лягушки в слабый раствор кислоты. Следовательно, торможение центров спинного мозга явилось следствием возбуждения центров среднего мозга. Прекращение этого тормозного контроля при перерыве цереброспинальных проводящих путей вызывает резкое повышение возбудимости спинальных центров и гиперрефлексию. - Принцип обратной афферентации - заключается в рецепторном восприятии результатов рефлекторного акта и проведении информации назад в структуры нервного центра, где она обрабатывается и сравнивается с сохраняющимися параметрами возбуждения. Обратная афферентация реализуется в виде положительной или отрицательной обратной связи. Таким образом, с помощью обратной афферентации нервные центры осуществляют непрерывный контроль эффективности, целесообразности и оптимальности рефлекторной деятельности.
Билет 18 Понятие о физиологической системе. Стратегия регулирования биологической системы. Основные формы приспособления организма к окружающей среде. Местная, гуморальная и нервная регуляция. Определение рефлекса. Рефлекторная дуга. Под физиологической системой понимают наследственно закрепленную, регулируемую систему органов и тканей (кровообращения, дыхания, пищеварения и т. д.), которые функционируют в организме не изолировано, а во взаимодействии друг с другом. Биологическое регулирование проявляется в том, что основным свойством живых систем с точки зрения их организации является способность упорядочивать свои связи со средой за счет поддержания равновесия и целостности биологической особи в процессе постоянного обмена с окружением веществом, энергией, информацией. Регулятивную значимость биологического как объективного регулирования обозначал еще В.И. Вернадский. «Жизнь, - писал он, - не является... внешним случайным явлением на Земной поверхности. Она теснейшим образом связана со строением земной коры, входит в ее механизм и в этом механизме исполняет величайшей важности функции, без которых он не мог бы существовать». Таким путем в природе формируются замкнутые циклы регуляции вещества, энергии, информации. Биологическое регулирование, противодействуя негативному воздействию Среды, состоит в активном стремлении биосистем к самосохранению себя как живого, в целях максимальной адаптации к условиям существования. Это - стратегическая цель биологической регуляции. С изменением внешних условий система самоизменяется, что определяет целенаправленность ее поведения. По отношению к биологическим системам целевая ориентация может быть определена как «модель потребного результата», возникающая на основе «опережающего отражения действительности». Принципиальной особенностью регулирования в биологических системах является регулирование по принципу автоупорядочивания. Автоупорядочивание как принцип появляется в биологических организациях и означает свойство систем собственными усилиями устанавливать и поддерживать на определяемом жизненной (необходимость выживания) или целевой (биологическая особь может действовать и вопреки жизненным установкам за счет проявления воли и терпения) ориентацией уровне те или иные параметры своего функционирования. При таком типе регуляторы воздействуют на систему не извне, а изнутри, они выработаны самой системой и непосредственно встроены в процессы ее функционирования. Биологическое регулирование проявляется в относительно объективной способности живых систем сохранять и передавать из поколения в поколение выработанные способы авторегулирования в виде определенных стереотипных моделей обеспечения выживания биологических особей. Передача регулятивной информации, в основном, осуществляется объективно наследственным генетическим путем, но на уровне высших животных присутствуют и негенетические механизмы передачи - научение, подражание, заражение и другие психические способы трансляции, которые уже можно отнести к субъективному социальному регулированию. Биологическое регулирование, таким образом, представляет собой процесс упорядочения активности биосистем, состоящей в стабилизации взаимосвязей со средой и структуры внутренних процессов, а также передачи регулятивной информации последующим поколениям в объективном или субъективном виде. Нервная регуляция обеспечивает быструю и направленную передачу сигналов, которые в виде нервных импульсов по соответствующим нервным проводникам поступают к определенному адресату — объекту регуляции. Быстрая передача сигналов (до 80-120 м/с) без затухания и потери энергии обусловлена свойствами проводящих возбуждение структур, преимущественно состоянием их мембран. Нервной регуляции подлежат как соматические (деятельность скелетной мускулатуры), так и вегетативные (деятельность внутренних органов) функции. Это универсальное значение нервной регуляции жизнедеятельности и физиологических функций было положено в основу концепции нервизма, рассматривающей целостность организма как результат деятельности нервной системы. Однако абсолютизация этой концепции до теории физиологии не оставляет места для многообразия уровней и связей в системе регуляции жизнедеятельности механизмов интеграции функций. Элементарный и основной принцип нервной регуляции — рефлекс. Гуморальная регуляция представляет собой способ передачи регулирующей информации к эффекторам через жидкую внутреннюю среду организма с помощью молекул химических веществ, выделяемых клетками или специализированными тканями и органами. Этот вид регуляции жизнедеятельности может обеспечивать как относительно автономный местный обмен информацией об особенностях метаболизма и функции клеток и тканей, так и системный эфферентный канал информационной связи, находящийся в большей или меньшей зависимости от нервных процессов восприятия и переработки информации о состоянии внешней и внутренней среды. Соответственно, гуморальную регуляцию подразделяют на местную малоспециализированную саморегуляцию, и высокоспециализированную систему гормональной регуляции, обеспечивающую генерализованные эффекты с помощью гормонов. Местная гуморальная регуляция (тканевая саморегуляция) практически не управляется нервной системой, тогда как система гормональной регуляции составляет часть единой нейрогуморальной системы. Нервная регуляция осуществляется с помощью электрических импульсов, идущих по нервным клеткам. По сравнению с гуморальной она: - происходит быстрее - более точная - требует больших затрат энергии - более эволюционно молодая. Гуморальная регуляция процессов жизнедеятельности осуществляется за счет веществ, выделяемых во внутреннюю среду организма (лимфу, кровь, тканевую жидкость). Гуморальная регуляция может осуществляться с помощью: - гормонов – биологически активных (действующих в очень маленькой концентрации) веществ, выделяемых в кровь железами внутренней секреции; - других веществ. Например, углекислый газ - вызывает местное расширение капилляров, к этому месту притекает больше крови; - возбуждает дыхательный центр продолговатого мозга, дыхание усиливается. Рефлекс - это ответная реакция организма на раздражение рецепторов, осуществляемая с помощью возбуждения нервной системы и имеющая приспособительное значение. В этом определении содержится сразу несколько важных признаков рефлекса: - это ответная реакция, а не самопроизвольная, - необходимо раздражение, без которого рефлекс не возникает, - в ос
|
||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-09-18; просмотров: 548; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.220 (0.014 с.) |