
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Созревание зернистых лейкоцитовСодержание книги
Поиск на нашем сайте
2-5 ядрышек, узкий ободок цитоплазмы, не со- держащий гранул (диаметр 12-20 мкм).
собная к делению клетка, в ядре отсутствуют яд- рышки.
Метамиелоцит (диаметр 12-15 мкм) имеет бухто- образное вдавление ядра, цитоплазма содержит нежную специфическую зернистость
В палочкоядерных лейкоцитах ядро имеет форму изогнутой палочки.
Сегментоядерные гранулоциты – это зрелые клет- ки (VI класс), ядро которых состоит из 2-4 сег- ментов.
Рис. 43. Стадии развития зернистых лейкоцитов
Материнские клетки миелобласты встречаются в норме в небольшом количестве в костном мозге (в 0,5-2% от всех ядер- ных клеток). Миелобласт представляет собой большую клетку (диаметром 10-20 мкм) с большим слабо окрашивающимся ядром и небольшой площадью цитоплазмы. Структура хроматина неж- но-сетчатая, с равномерной окраской и калибром нитей. Ядра мо- гут содержать от 2 до 5 мелких голубых ядрышек. Цитоплазма голубая («чистое голубое небо») и не содержит зернистости. Дальнейшее развитие миелобласта приводит к переходу его в миелоцит, который является преобладающей формой гранулоци- тов в костном мозге. Между миелобластом и миелоцитом имеется ряд переходных форм, которые объединены под названием про- миелоцитов, которые в костном мозге встречаются в 3 раза чаще, чем миелобласты. В последних содержится много азурофильной зернистости (самые «грязные» клетки). Зернистость содержит ка- тионные белки хром-содержащий энзим миелопероксидазу. Клетки V класса (созревающие) проходят через различное количество стадий. В процессе созревания гранулоцитов (ней- трофилов, эозинофилов и базофилов) их ядро уплотняется и под- вергается сегментации. В цитоплазме при окраске по Райту появ- ляются специфические нейтро-, эозино- или базофильные грану- лы. Созревание гранулоцитов заключается в уменьшении его размеров и в изменениях тонкой структуры цитоплазмы и ядра. Изменения в цитоплазме заключаются в постепенном уменьше- нии содержания в ней базофильного вещества. Поэтому цвет ци- топлазмы при окраске по Романовскому-Гимза постепенно пере- ходит из синего в светло-голубой. Одновременно с ослаблением базофилии в цитоплазме начинает появляться зернистость. Сна- чала зернистость азурофильна, но по мере дальнейшего созрева- ния она становится дифференцированной, приобретает окраску, которая свойственна зернистости вполне сформировавшегося миелоцита. Одновременно с изменениями в цитоплазме происходят и изменения в ядре: последнее уменьшается, приобретает круглую или овальную пузырькообразную форму, хроматин располагается в нем в виде толстых перекладин. В миелобластах и промиелоци- тах по характеру зернистости нельзя еще узнать черты тех трех видов гранулоцитов, которые из них развиваются в дальнейшем. Дифференциация клеток гранулоцитарного ряда на нейтрофилы, эозинофилы и базофилы начинается с миелоцита. Нейтрофильная зернистость мелкая, сине-фиолетовая, пылевидная, эозинофиль- ная – крупная, желто-розово-оранжевого цвета, одинакового раз- мера («красная икра»), базофильная – разнокалиберная (от очень крупных до пылевидных), темно-фиолетового цвета. В костном мозге можно встретить три вида миелоцитов: нейтрофильные, эоэинофильные и базофильные. Каждый вид миелоцита дает при дальнейшем развитии соответственный зрелый гранулоцит: из нейтрофильного миелоцита развивается нейтрофильный лейко- цит, из эозинофильного – эозинофильный, а из базофильного – базофильный. Созревание миелоцита при переходе в зрелый гранулоцит заключается в уменьшении размеров и в изменении ядра. По- следнее приобретает более сложную структуру, становится сна- чала почкообразным (у метамиелоцитов), затем подковообраз- ным (у палочкоядерных лейкоцитов), начинает разделяться на отдельные лопасти и, наконец, на отдельные сегменты, соеди- ненные между собой тоненькими мостиками. Эта сегментация слабо выражена в базофилах, более выражена в эозинофилах и достигает своего, окончательного развития в нейтрофилах. Структура ядра миелоцита зависит от степени зрелости: молодые формы имеют ядра более равномерного (рыхлого) строения, бо- лее зрелые – компактную, глыбчатую структуру. Таким образом, признаками незрелости («юности») гранулоцитов являются большие размеры клетки, недифференцированное и бледно ок- рашенное ядро, базофилия цитоплазмы и отсутствие в ней зерни- стости (за исключением миелоцитов). Зрелые нейтрофилы (VI класс) представляют собой круглые клетки в 1,5-2 раза больше эритроцита; при окраске по методу Романовского-Гимза цитоплазма нейтрофилов имеет слабо- розовую окраску. Ядра нейтрофилов густо окрашиваются основ- ной краской в сине-фиолетовый цвет. Ядро состоит из больших петель хроматина, между которыми находится более светло ок- рашенный парахроматин. В зрелых нейтрофилах ядра состоят из 2-4 сегментов, соединенных между собой тоненькими мостиками (сегментоядерные нейтрофилы). В цитоплазме отмечается тонкая пылевидная зернистость красновато-фиолетового цвета. В нормальной крови встречаются в небольшом количестве нейтрофилы менее зрелые; ядро таких клеточных элементов не сегментировано, а представляется в виде узкого вытянутого кол- басовидного образования (палочкоядерные нейтрофилы). В очень редких случаях в нормальной периферической крови могут попа- даться еще более молодые нейтрофилы с компактным ядром, имеющим бухтообразное вдавление, которое придает ядру поч- ковидную форму – метамиелоциты (юные). Время созревания нейтрофилов в костном мозге составляет 10-14 дней. При инфекционно-воспалительных процессах и большей, чем в норме, потребности в нейтрофилах, этот процесс значительно ускоряется. Палочкоядерные и сегментоядерные нейтрофилы не покидают сразу костный мозг, остаются в нем в течение 3-4-х дней, формируя костно-мозговой резерв нейтрофи- лов, которые каждые 3-5 дней обновляются. В периферической крови выделяют два пула лейкоцитов: циркулирующий и маргинальный (краевой), количества их прак- тически равны. Время циркуляции нейтрофилов в сосудистом русле равно 4-6 часам. Из сосудистого русла нейтрофилы через стенку сосудов выходят в ткани, где участвуют в защитных им- мунных реакциях. Выполнив свою функцию в тканях, нейтрофи- лы разрушаются путем апоптоза. Эозинофилы несколько больше по размерам, чем нейтрофи- лы. Ядро их состоит обычно из двух грушевидных, соединенных тоненьким мостиком лопастей. Для эозинофилов характерна обильная грубая зернистость, густо рассеянная в протоплазме и окрашивающаяся эозином в ярко-красный цвет. Эозинофилы имеют ферменты: гистаминазу, арилсульфата- зу, благодаря чему являются инактиваторами гистамина, гепари- на, лейкотриенов, тромбоксанов, простагландинов и поэтому уча- ствуют в воспалительных процессах и, особенно, при аллергиче- ских заболеваниях, в патогенезе которых существенное место принадлежит гистамину. Этим определяется важная функция эо- зинофилов, которую они выполняют в здоровом организме: ней- трализация избытка гистамина и других подобных биологически активных веществ, поддержание на нормальном уровне микро- циркуляции в тканях. Увеличениесодержания эозинофилов в крови при воспале- нии образно называют «алой зарей выздоровления». Также в эо- зинофилах образуется ряд неферментативных катионных белков, которые могут разрушать оболочки гельминтов. Эозинофилы созревают в костном мозге за 3-5 дней. Не об- разуя резерва, выходят в сосудистое русло, где циркулируют в течение 5-24 часов. Тканевые эозинофилы могут возвращаться в сосудистое русло, чем объясняют длительность эозинофильных реакций. Базофилы несколько меньше нейтрофилов. Протоплазма со- держит многочисленные крупные зерна, окрашенные основной краской в сине-фиолетовый цвет. Эта зернистость покрывает большую часть ядра. Ядро неправильной формы, лопастное, ино- гда разделено на две или три части. Базофилы содержат в своих гранулах гистамин, гепарин, се- ротонин и некоторые другие биологически активные вещества. В крови их содержание крайне низкое, а аналоги базофилов – туч- ные клетки находятся по периферии вдоль стенок кровеносных сосудов, в подслизистых слоях желудочно-кишечного, респира- торного тракта, в плевре и брюшине. После фиксации на них им- мунных комплексов, включающих антиген и две молекулы IgE, происходит высвобождение медиаторов воспаления (аллергии) тучными клетками. Поэтому им принадлежит большая роль в па- тогенезе аллергических реакций I типа (реагиновый или анафи- лактический тип реакций). Базофилы принимают участие в образовании и накоплении гепарина, транспортируют гепарин к стенке сосудов, благодаря чему участвуют в процессах свертывания крови и метаболизме триглицеридов. Базофилы также участвуют в обмене гистамина, в них содержится половина всего гистамина крови. В ответ на по- падание в организм аллергена и образование комплекса антиген- антитело происходит дегрануляция базофилов с выбросом гиста- мина и других биологически активных веществ. Гранулоциты (нейтрофилы, эозинофилы и базофилы) влия- ют на развитие сосудистых реакций при воспалении, стимулиру- ют процессы тканевой регенерации. Лимфоциты по происхождению представляют собой клетки лимфоидного ряда. Место их образования – лимфоидная ткань лимфатических узлов, селезенка и отчасти костный мозг, откуда они поступают в кровь. Лимфоциты – нефагоцитирующие лейкоциты, они осущест- вляют и регулируют иммунный ответ. Различают две основные разновидности лимфоцитов: Т-лимфоциты (тимус-зависимые) – составляют основную часть (70%) циркулирующих лимфоцитов и В-лимфоциты (бурса-зависимые), а также менее многочислен- ный класс естественных, или «натуральных», нормальных килле- ров. На мембранах лимфоцитов различных стадий развития при- сутствуют поверхностные антигены лимфоцитов (кластеры диф- ференцировки, или СD-антигены «сluster differentiation». Последние выявляются в реакциях с моноклональными ан- тителами. Среди Т-лимфоцитов, в зависимости от экспрессируе- мых ими рецепторов CD, выделяют следующие популяции: Т- хелперы (CD4), цитотоксические Т-лимфоциты и Т-супрессоры (CD8). Образовавшись из стволовых клеток костного мозга, не- зрелые протимоциты «заселяют» корковый слой тимуса, а затем мигрируют в селезенку, где созревают и приобретают характер- ные для Т-лимфоцитов антигенные свойства. Одной из основных функций лимфоцитов в организме явля- ется участие их в иммунных процессах. Для осуществления этих функций различные клоны лимфоцитов способны к синтезу уни- кальных белковых распознающих молекул, которые находятся на их поверхности, либо секретируются и циркулируют в биологи- ческих жидкостях организма, как продукты плазматических кле- ток – иммуноглобулины. Т-лимфоциты играют ключевую роль в клеточном иммунитете, который обеспечивает уничтожение чу- жеродных клеток, в том числе атипичных. Т-лимфоциты опосре- дуют аллергические реакции замедленного типа, отторжение ал- лотрансплантата, реакцию «трансплантант против хозяина», кон- тактную аллергию, иммунитет против опухолей, вирусов, гриб- ков и внутриклеточных паразитов. Способность T-лимфоцитов участвовать в специфическом иммунном ответе (на определен- ный антиген) обусловлена наличием вариабельной части рецеп- тора, возможности изменчивости ее структуры и селективным размножением лимфоцитов сенсибилизированных именно к дан- ному антигену. В-лимфоциты составляют до 20% циркулирующих лимфо- цитов. В-лимфоциты, превращаясь в плазмоциты, участвуют в реализации всех видов гиперчувствительности немедленного ти- па посредством образования иммуноглобулинов (Ig G, M, A, D, E) – нейтрализации токсинов и вирусов, опсонизации бактерий для фагоцитоза, антителозависимой клеточной цитотоксичности. Лимфоциты, наряду с клетками системы мононуклеарных фаго- цитов и плазматическими клетками, принимают участие в обра- зовании антител. В-лимфоциты находятся в основном в гермина- тивных центрах коркового вещества лимфатических узлов и в бе- лой пульпе селезенки. Они экспрессируют поверхностные анти- гены CD19, CD20 и CD22. При взаимодействии с антигеном (чу- жеродным белком) лимфоциты не только разрушают его, но и со- храняют информацию о свойстве этого антигена, т. е. являются хранителями «иммунной памяти», обеспечивающей ответ на по- вторное введение этого же антигена. В-лимфоциты могут жить в течение многих лет. Нормальные (натуральные киллеры, NK-клетки) – это большие лимфоциты, способные непосредственно, без помощи антител, уничтожать клетки-мишени (опухолевые и инфициро- ванные вирусами или бактериями клетки). Они имеют некоторые антигены Т-лимфоцитов и макрофагов, а также характерные только для этих клеток CD16. Созреваниелимфоцитов. Материнская клетка – лимфобласт обнаруживается в зародышевых центрах лимфатических узлов и фолликулов селезенки. Это большая клетка с базофильно окра- шивающейся по методу Романовского-Гимза в голубой цвет ци- топлазмой, большим, бледно окрашенным ядром, в котором со- держатся 1-2 светло-голубых ядрышка. Структура хроматина в ядре мелкогранулярная, равномерная. По мере созревания клетки размеры ее уменьшаются, ядро становится более компактным и окрашивается в более темный фиолетово-красный цвет.
Рис. 44. Стадии развития лимфоцитов Зрелые лимфоциты равны или несколько больше эритроци- тов, имеют диаметр от 7 до 15 мкм. Цитоплазма при окраске по методу Романовского-Гимза окрашена в светло-голубой цвет. Ядро круглой или слегка овальной формы окрашивается основ- ной краской в голубой цвет. Оно часто имеет с одной стороны бухтообразное вдавление. Вокруг ядра остается бледный ободок, более светлый, чем остальная цитоплазма (перинуклеарная зона). В цитоплазме отдельных лимфоцитов можно иногда при хорошей окраске обнаружить несколько красновато-фиолетовых крупных гранул (азурофильная зернистость). Отличить на окрашенном препарате различные стадии раз- вития лимфоцита (лимфобласт, пролимфоцит, лимфоцит) трудно, так как они сходны между собой. Лимфобласты также с большим трудом удается отличить от миелобластов. Однако отличитель- ным признаком является положительная реакция на оксидазу и пероксидазу у миелобластов и отрицательная – у лимфобластов. Моноциты входят в систему мононуклеарных фагоцитов, элементы которой рассеяны во многих местах организма, в том числе в костном мозге. На поверхности моноцитов и макрофагов имеется множест- во рецепторов: для иммуноглобулинов, различных антигенов, белков системы комплемента, лимфокинов, ряда гормонов и др. В цитоплазме находятся гранулы (лизосомы), содержащие фер- менты. Макрофаги выполняют важные функции: участие в не- специфическом и специфическом иммунном ответе вместе с Т- лимфоцитами в качестве антигенпрезентирующих клеток, уча- стие в обмене веществ (метаболизм липидов, железа, пигментов). Моноциты образуются в костном мозге, где проходят сле- дующие морфологически различимые стадии созревания: моно- бласт, промоноцит, моноцит. Моноциты крови не являются еще окончательно зрелыми клетками – в тканях они превращаются в макрофаги. Ранние предшественники моноцитов развиваются из поли- потентной стволовой клетки костного мозга под влиянием гемо- поэтинов. Дифференцировка моноцитов из монобластов проис- ходит в течение 5 дней, после чего они сразу выходят в кровоток, не образуя костномозгового резерва. Моноциты циркулируют в
монобласт промоноцит моноциты
Рис. 45. Стадии развития моноцитов
крови от 30 до 104 часов и затем уходят в ткани, где под влияни- ем различных тканевых факторов дифференцируются в органо- и тканеспецифические макрофаги. В результате образуются много- ядерные клетки инородных тел, остеокласты, эпителиоидные клетки. Продолжительность жизни тканевых макрофагов от не- скольких часов до нескольких месяцев и лет. Монобласт имеет округлое либо округло-вытянутое ядро, иногда бобовидное, нежной структуры с ядрышками и напомина- ет миелобласт. У промоноцита имеется изогнутое ядро с волни- стыми очертаниями. Цитоплазма окрашивается в базофильные тона. Моноциты самые большие клетки нормальной перифериче- ской крови: их диаметр колеблется от 14 до 20 мкм. Цитоплазма моноцитов при окраске по методу Романовского-Гимза серо- голубого цвета. Ядро крупное, неправильно-круглой, овальной, почковидной или подковообразной формы, расположено в клетке эксцентрично. Оно бедно хроматином и потому слабо окрашено, так что между ним и цитоплазмой нет такого контраста, как в других лейкоцитах. В отличие от лимфоцитов в моноцитах пери- нуклеарной зоны нет. При очень хорошей окраске в цитоплазме обнаруживается чрезвычайно мелкая азурофильная зернистость. Содержание лейкоцитов у взрослых составляет (4-9) х 109 в
Рис. 46. Изменение процентного содержания нейтрофилов (N) и лимфоцитов (L) в крови у детей в зависимости от возраста по Туру, Шабалову.
«Правило четырех четверок» – относительное содержание лимфоцитов и нейтрофилов уравнивается в 4 года на уровне около 44 %.
Затем процент лимфоцитов повышается, а нейтрофилов – снижается, к 4 суткам их количество уравнивается (первый пере- крест). К 1-2 годам процентное содержание лимфоцитов достига- ет 65%, а нейтрофилов – 25%. Затем идет повторное снижение лимфоцитов и возрастание нейтрофилов. У детей с 5-6 дня до 5-6 лет в лейкоцитарной формуле крови преобладают лимфоциты над нейтрофилами. К 5-6 годам показатели содержания нейтро- филов и лимфоцитов выравниваются, достигая в среднем 44% (2- й перекрест). К 5-му году жизни общее содержание лейкоцитов составляет около 10 × 109/л и лишь с 10-го года становится равным числу лейкоцитов крови у взрослого. К 14-15 годам лей- коцитарная формула не отличается от таковой у взрослых Количество лейкоцитов в крови колеблется также в течение дня, достигая максимума в вечерние часы. Подсчета количества форменных элементов крови недоста- точно для суждения о характере нарушений в организме. Необ- ходимо также детальное ознакомление с качественными измене- ниями отдельных клеток крови и с количеством отдельных видов лейкоцитов. Это становится возможным при микроскопическом исследовании окрашенных по Романовскому-Гимза мазков крови и подсчете лейкоцитарной формулы.
Лейкоцитарная формула (относительная лейкоцитарная формула, или лейкограмма) – это процентное содержание от- дельных видов лейкоцитов в крови.
Лейкоцитарный профиль (абсолютная лейкоцитарная формула) отражает абсолютное количество лейкоцитов каждого вида в единице объема крови.
Диагностическое значение изменения процентного соот- ношения форменных элементов в лейкоцитарной формуле. Следует подчеркнуть, что изменение процента какого-либо вида лейкоцитов в лейкоцитарной формуле по сравнению с нормаль- ной, не всегда означает такое же изменение абсолютного числа данных лейкоцитов в крови. Так, например, если при нормальном количестве в 8000 лей- коцитов в 1 мкл крови процент лимфоцитов составляет 25, то аб- солютное количество их в 1 мкл крови равно 2000. Если же при общем количестве лейкоцитов в 2000 в 1 мкл крови лимфоцитов содержится 50 %, т. е. в два раза больше нормального, то абсо- лютное количество лимфоцитов в 1 мкл крови составляет 1000 (ниже нормы), т.е. имеет место лимфопения, а такое увеличение процента лимфоцитов в лейкоцитарной формуле рассматривается как относительный лимфоцитоз. В некоторых случаях важно учитывать не только лейкоци- тарную формулу, но и миелограмму (приложение 2, 3), представ- ляющую собой процентное соотношение различных клеток кост- ного мозга, находящихся на разных стадиях созревания.
Изменения
Примечание: в скобках указаны абсолютные цифры (760) (950) (1900) (190) сдвигом влево
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-15; просмотров: 783; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.5 (0.015 с.) |

 Миелобласты имеют большое круглое ядро с нежной сетчатой структурой хроматина, а также
Миелобласты имеют большое круглое ядро с нежной сетчатой структурой хроматина, а также Миелоцит (диаметр 14-16 мкм) – последняя спо-
Миелоцит (диаметр 14-16 мкм) – последняя спо- Лимфобласты (диаметр 20-22 мкм) имеют четкую перинуклеарную зону, грубую структуру хрома- тина и 1-2 ядрышка.
Лимфобласты (диаметр 20-22 мкм) имеют четкую перинуклеарную зону, грубую структуру хрома- тина и 1-2 ядрышка. Пролимфоцит (диаметр 11-12 мкм) с ядром рых- лой структуры, наличием ядрышка либо его ос- татков, наличием ободка цитоплазмы различной ширины и азурофильной зернистостью в T- лимфоцитах.
Пролимфоцит (диаметр 11-12 мкм) с ядром рых- лой структуры, наличием ядрышка либо его ос- татков, наличием ободка цитоплазмы различной ширины и азурофильной зернистостью в T- лимфоцитах. Лимфоцит (диаметр 7-15 мкм) с компактным ядром, глыбчатой структуры с небольшими про- светлениями и узким либо широким ободком ба- зофильной, либо бесцветной цитоплазмы.
Лимфоцит (диаметр 7-15 мкм) с компактным ядром, глыбчатой структуры с небольшими про- светлениями и узким либо широким ободком ба- зофильной, либо бесцветной цитоплазмы.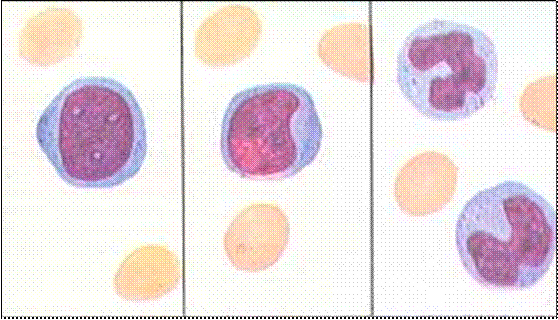
 1 л. В течение 1-х суток после рождения общее содержание лей- коцитов может достигать (10-30) х 109/л, а в течение первых двух недель – (5-20) х 109/л крови. Соотношение нейтрофилов и лим- фоцитов при этом, такое же, как и у взрослых (рис. 46).
1 л. В течение 1-х суток после рождения общее содержание лей- коцитов может достигать (10-30) х 109/л, а в течение первых двух недель – (5-20) х 109/л крови. Соотношение нейтрофилов и лим- фоцитов при этом, такое же, как и у взрослых (рис. 46).
 Таблица 11. Лейкоцитарная формула и лейкоцитарный профиль в норме и патологии
Таблица 11. Лейкоцитарная формула и лейкоцитарный профиль в норме и патологии



 Нейтрофилы
Нейтрофилы в гемограмме
в гемограмме



