Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Принцип конкурентного исключения. Экологическая диверсификацияСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
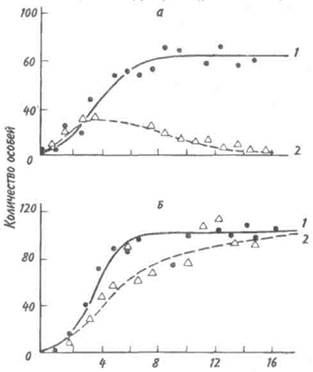
Решающее значение в природе имеет межвидовая конкуренция, поскольку она в большей степени, чем другие гетеротипические реакции, определяет роль видов в экосистемах. Межвидовая конкуренция — это такое взаимодействие, когда два вида соперничают из-за одних и тех же источников существования — пищи, пространства и т. п. Причем она возникает в тех случаях, если использование источника ресурсов одним видом ведет к ограничению использования его другим. Внутривидовая конкуренция сильнее межвидовой, но правило конкуренции распространяется и на последнюю: конкуренция между двумя видами тем сильнее, чем бли- же их потребности. В крайнем случае, можно допустить, что два вида с совершенно одинаковыми потребностями не могут существовать вместе: один из них через какое-то время обязательно будет вытеснен. Это положение получило статус закона, известного под названием принципа конкурентного вытеснения, или принципа Гаузе — в честь Г. Ф. Гаузе, впервые продемонстрировавшего это явление в экспериментах с инфузориями (рис. 6.2). При изолированном содержании численность популяций обоих видов инфузорий (рис. 6.2, кривые 1)
Рис. 6.2. Конкурентное исключение, продемонстрированное в лабораторном эксперименте на двух видах инфузорий (по Гаузе, 1934). a —Paramecium caudatum, б —P. aurelia: 1 — в изолированной культуре, 2 —в смешанной культуре. увеличивается до тех пор, пока не достигнет максимума. В смешанной культуре популяция второго вида (рис. 6.2,б, кривая 2) растет быстрее, чем первого (рис. 6.2,а, кривая 2), что приводит к ограничению ее численности. Если не произойдет изменений в. параметрах конкуренции, то конкурентное исключение (вымирание первой популяции)—лишь вопрос времени. Иногда два близких вида, имеющих одинаковые пищевые потребности, живут на одной территории, не конкурируя друг с другом. Такие, казалось бы, исключения из принципа Гаузе можно обнаружить среди птиц. В Англии большой баклан и хохлатый баклан совместно гнездятся на одних и тех же скалах и кормятся в одних и тех же водах, но они вылавливают разную пищу. Большой баклан ныряет глубоко и питается преимущественно бентическими животными (камбала, креветки), тогда как хохлатый баклан охотится в поверхностных водах на сельдевых рыб. Подобные факты, а их много, подводят нас к понятию экологической ниши, предложенному Элтоном еще в 1927 г. По образному выражению Одума, местообитание — это адрес вида, тогда как экологическая ниша — это род его занятий. Иными словами, знание экологической ниши дает возможность ответить на вопросы: где, чем и как питается вид, чьей добычей он сам является, каким образом и где он размножается и т. д. Современное определение звучит следующим образом: экологическая ниша — это роль вида в переносе энергии в экосистеме. Имеющиеся в экологической литературе трактовки данного понятия чрезвычайно разнообразны. Все же можно дать следующую наиболее общую формулировку: экологическая ниша — это общая сумма всех требований организма к условиям существования, включая занимаемое им пространство, функциональную роль в сообществе (например, трофический статус) и его толерантность по отношению к факторам среды — температуре, влажности, кислотности, составу почвы и др. Названные три критерия определения экологической ниши удобно обозначать как пространственную, трофическую и многомерную ниши. Пространственная ниша, или ниша местообитания, может быть названа «адресом» организма. Трофическая ниша характеризует особенности питания и, следовательно, роль организма в сообществе, как бы его «профессию». Многомерная, или гиперпространственная, ниша — это диапазон всех условий, при которых живет и воспроизводит себя особь или популяция. Для интерпретации этого понятия используется теория множеств. Многомерная ниша — это область в гиперпространстве, измерениями которого служат различные экологические факторы; она охватывает диапазоны толерантности по каждому фактору-измерению. Такой акцент исключает из рассмотрения функциональный и поведенческий аспекты ниши. Различают фундаментальную (потенциальную) нишу, которую организм мог бы занимать в отсутствие конкурентов, хищников и других врагов и в которой физические условия оптимальны, и реализованную нишу — фактический диапазон условий существования организма, который или меньше фундаментальной ниши, или равен ей. Фундаментальную нишу называют иногда преконкурентной, а реализованную — постконкурентной. Однако это не синонимы, поскольку не только конкуренция ограничивает ширину реализованной ниши. Взаимоотношение между фундаментальной и реализованной нишами может быть пояснено с помощью так называемой лицензионной модели. Понятие «экологической лицензии» было впервые введено Гюнтером, но оно было весьма расплывчатым и определялось как условия внешней и внутренней среды, разрешающие осуществляться некоторым эволюционным факторам и событиям. Позднее В. Ф. Левченко и Я. И. Старобогатов вернулись к понятию экологической лицензии, причем были учтены следующие особенности: место в пространстве и времени, роль потоков вещества и энергии и наличие градиентов внешних условий, обеспечиваемых экосистемой для популяций и организмов там существующих. Приведенное описание лицензии напоминает те слова, которые используют, когда говорят о так называемой «пустой экологической нише». Однако, строго говоря, последняя не может быть объяснена, если использовать классические определения ниши. Любая популяция имеет фундаментальную (потенциально возможную) и реализованную (фактическую) экологическую нишу (рис. 6.3). Реализованная ниша RN никогда не выходит за границы лицензии L, но при этом обязательно перекрывает фундаментальную нишу FN. Если внутри лицензий экосистемы находится по одной популяции, то мы имеем дело со случаем простой экоси-
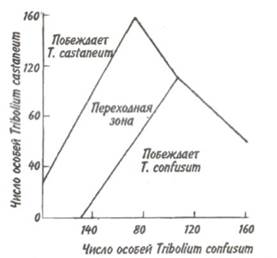
Рис. 6.3. Взаимодействия между фундаментальными нишами Видов (FN), реализованными нишами популяций (RN) и лицензиями (L.) в пространстве внешних факторов (Р1 и Р2). А-С — различные варианты взаимодействия ниш и лицензий. Реализованные и фундаментальные ниши различным образом затенены, зачерненные фрагменты — области конкуренции (по Левченко, 1993). стемы. При наличии в лицензии нескольких популяций между ними возможна конкуренция за ресурсы. В этом случае мы имеем случай сложной экосистемы. Для сложной экосистемы можно говорить о том, что у нее имеются групповые фундаментальные ниши, а в лицензии каждого трофического уровня существуют групповые реализованные ниши. Теперь конкретизируем принцип Гаузе: два вида не могут занимать одну и ту же экологическую нишу. Принцип Гаузе явился важным экологическим обобщением. Он позволил увидеть, что для сосуществования видов в конкурентных сообществах абсолютно необходимы какие-то различия их экологических ниш. Поскольку для каждой из конкурирующих сторон избегание взаимодействия выгодно, конкуренция играет роль мощного эволюционного фактора, приводящего к разделению ниш, специализации видов и возникновению видового разнообразия. Явление разделения экологических ниш в результате межвидовой конкуренции получило название экологической диверсификации. Экологическая диверсификация между обитающими совместно (т.е. симпатрическими) видами в большинстве случаев осуществляется по следующим трем параметрам: пространственному размещению, пищевому рациону и распределению активности во времени. Как правило, для значительного ослабления и даже полного устранения конкуренции достаточно возникновения четких различий по одному из этих параметров. Различия в пространственном размещении заключаются в выборе видами разных стаций или даже микростаций. Большой пестрый дятел ищет пищу в основном на стволах деревьев, средний пестрый — на больших ветвях, а малый пестрый дятел — на ветках кроны. Благодаря пути из варяг в греки на север проникла черная крыса. Она загнала серую крысу в подполье, а сама обосновалась на чердаках. Когда в связи с развитием судоходства на юге появилась серая крыса, или пасюк, то она вытеснила черную крысу на чердачные помещения, а сама обосновалась в подполье и канализационной сети. Экологическая диверсификация по пищевому рациону— широко распространенное явление. Не раз было показано, что близкородственные виды животных характеризуются различными пищевыми потребностями. Пищевые рационы пустынных ящериц, принадлежащих к нескольким симпатрическим видам, состоят у одних преимущественно из муравьев, у других из термитов, у третьих из ящериц других видов или только растений. Сходные примеры различий в рационах у родственных симпатрических видов известны для многих птиц и млекопитающих. Разделение ниш часто бывает сопряжено с распределением активности видов во времени. Примерами могут служить разные типы суточной (ласточки и летучие мыши) или сезонной (некоторые виды ящериц, насекомых) активности. В умеренных широтах у садовой и черноголовой славок наблюдается сдвиг в периодах кладки в две недели. В тропиках эти виды могут размножаться круглый год, так как постоянное обилие необходимых для их питания насекомых ослабляет конкуренцию. Конкуренция за пищу приводит иногда к ускорению развития и изменению всего жизненного цикла вида. Интересный пример такой конкуренции представляют муха-саркофагида и оса-сфекс. Саркофагида заражает саранчовое, погружая яйцо в его тело. Сфекс парализует ослабленное саранчовое, после чего затаскивает его, а заодно и яйцо саркофагиды, в свою норку/ Обычно личинка паразитической мухи некоторое время развивается, но затем погибает, не выдержав пищевой конкуренции с быстро растущей личинкой сфекса. Однако отдельные личинки саркофагид благодаря ускорению развития успевали съесть парализованное саранчовое до появления личинки осы. Поскольку личинки саркофагид попадают в норку сфекса более или менее регулярно, то в эволюции закрепилась более высокая скорость развития личинок саркофагид, следствием чего стала меньшая плодовитость мух. Одним из следствий экологической диверсификации является так называемое смещение признаков. Иногда особи двух широко распространенных видов более сходны в тех частях своих ареалов, в которых встречаются по отдельности (аллопатрически), чем на участках совместного обитания. Например, два вида поползней столь похожи друг на друга в районах аллопатрии, что различить их может только специалист. Но в областях совместного обитания они имеют специфические признаки: у одного клюв и черная заглазничная полоса больше, чем у другого. Исторически возникшее приспособление к наиболее полному использованию условий местообитания и к пространственному расселению у животных и растений привело к образованию жизненных форм. Их группирование по тем или иным признакам являет собой примеры экологической классификации растительного и животного мира. Если систематическая классификация растений и животных строится на основе родства организмов, то экологическая классификация (т. е. классификация жизненных форм) выявляет возможные пути приспособления организмов к среде обитания. В качестве примера жизненных форм у растений можно привести травы, кустарнички, кустарники, деревья и т. д., а среди животных — плавающие формы, роющие формы и т. п. Жизненные формы организмов следует отличать от экологических групп организмов. Если первые отражают приспособленность организмов к специфике данного местообитания в целом, то экологические группы отражают приспособленность организмов к отдельным факторам внешней среды. Например, по отношению к условиям увлажнения выделяются экогруппы — мезофиты, ксерофиты, суккуленты и т. д., к условиям освещения — светолюбы и тенелюбы, по отношению к типам почв — криофиты (жители холодных почв), галофиты (солерсы) и т. д. Представители одной и той же жизненной формы могут принадлежать к разным экологическим группам. Например, клевер луговой и астрагал выделяющийся относятся к стержнекорневым растениям, но принадлежат к разным экологическим группам: клевер, произрастающий на лугах, является мезофитом, астрагал, обитающий в сухостепных районах,— ксерофитом. Манжетка обыкновенная и копытень европейский, будучи представителями короткокорневищных растений, по отношению к свету распределяются по разным экогруппам: манжетка является светолюбом, а копытень — тенелюбом. В то же время оба вида по отношению к фактору влажности принадлежат к одной экологической группе — мезофитам. На успехе в конкурентной борьбе того или иного вида сказывается влияние различных абиотических факторов. Воздействие среды на исход конкуренции изучалось в многочисленных лабораторных экспериментах с двумя видами малого мучного хрущака, Tribolium castaneum и T.confuswn. Было установлено, что при равенстве исходной численности жуков обоих видов в жарком влажном климате (t0 = 34°С, относительная влажность 70%) всегда побеждает Т.castaneum. При ином соотношении начальной численности популяций независимо от факторов среды успех сопутствует то одному, то другому виду (рис. 6.4). Первоначальное доминирование какого-либо вида увеличивает вероятность его победы. При незначительном перевесе в численности той или иной популяции (переходная зона) каждый из видов может одержать победу в зависимости от условий температуры и влажности.
Рис. 6.4. Зависимость исхода конкуренции между лабораторными линиями жуков Tribolium castaneum И Т. confusum от начальных плотностей особей (из Пианка, 1981). Оценивая успех в конкурентной борьбе одного из двух видов в природных условиях, опираться на такой показатель, как исходная плотность популяции, не представляется возможным. Как правило, вид более специализированный, стенобионтный, оказывается и более конкурентоспособным по сравнению с видом эврибионтным. Один из двух видов плоских червей обитает только в верховьях рек при температуре воды от 6,5 до 16 °С, а другой вид, выносящий температуры в пределах от 6,5 до 23 °С, может жить на всем протяжении реки. Когда оба вида симпатрические, то стенотермный вид вытесняет эвритермный из верховьев, и последний сохраняется только в нижнем течении реки, где значения температуры воды колеблются между 13 и 23 °С. На Кубе обитают два вида крокодилов: острорылый и кубинский. Несмотря на общие биологические особенности они, за редким исключением, никогда вместе не встречаются. Острорылый крокодил имеет большую экологическую пластичность, он хорошо переносит осолонение водоемов, поэтому широко распространен во многих прибрежных водоемах: устьях рек, лагунах и других низких местах. В отличие от острорылого кубинский крокодил встречается только в пресных водах. При совместном обитании он как более специализированный и поэтому более конкурентоспособный вытесняет острорылого крокодила из пресных водоемов, принуждая его переходить в солоноватые воды. Важным параметром вида является ширина его экологической ниши. Этот показатель всегда относителен. Мы можем говорить о широкой или узкой нише какого-нибудь организма лишь в сравнении с другими. Например, гусеницы дубовой листовертки, питающиеся только листьями дуба, имеют более узкую пищевую нишу, чем гусеницы непарного шелкопряда, развивающиеся на многих видах лиственных деревьев. Эврибионтные организмы обычно имеют более широкие ниши, чем стенобионтные. Однако экологическая ниша может иметь различную ширину по разным измерениям, таким как пространственное распределение, трофические связи и др. Когда два организма различных видов используют одни и те же ресурсы, их ниши перекрываются. Перекрывание может быть полным или частичным, может быть по одному измерению ниши, но не быть по другим. Если какие-то ресурсы имеются в недостаточном количестве, то при перекрывании ниш возникает конкуренция, которая тем интенсивнее, чем сильнее это перекрывание. На рис. 6.5 показаны разные варианты взаимодействия экологических ниш. В случае, когда ниши полностью разобщены (рис. 6.5, в), населяющие их виды не конкурируют друг с другом. Если они вообще не вступают в конкуренцию с другими организмами, то их ниши близки к потенциально возможным в данной ситуации. Когда экологические ниши частично совпадают (рис. 6.5,6), сосуществование видов становится возможным благодаря специфичности их приспособлений,
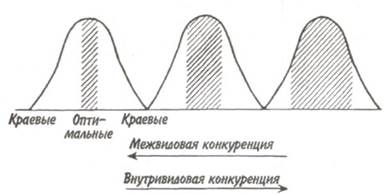
Рис. 6.5. Примеры возможных взаимодействий экологических ниш видов А и В (по: Пианка, 1981). Объяснения в тексте. например, особи одного вида занимают убежища, недоступные для более активного конкурента. Наконец, когда ниша одного вида включает в себя нишу второго вида или эти ниши полностью перекрываются (рис. 6.5,а), произойдет конкурентное исключение одного из видов или доминирующий конкурент оттеснит своего соперника на периферию зоны приспособленности. Реальная экологическая ниша слабого конкурента предельно сузится, заняв лишь некоторую часть потенциально возможной ниши. Именно поэтому целый ряд животных и растений живет в условиях, отличных от оптимальных. Например, многие растения, принимаемые за кальциефобные, переносят в чистой культуре самое различное содержание кальция; в природных условиях эти растения сдерживаются в границах своего ареала конкуренцией других видов. Конкуренция приводит к очень важным экологическим последствиям. В природе особи каждого вида являются объектами одновременно межвидовой и внутривидовой конкуренции. Межвидовая конкуренция оказывает прямо противоположное, по сравнению с внутривидовой, влияние на пределы выносливости популяции, способствуя сужению диапазона используемых ею местообитаний и ресурсов. Это происходит потому, что разные виды обычно имеют различную способность осваивать
Рис. 6.6. Влияние межвидовой и внутривидовой конкуренции на распределение местообитаний у птиц (из Одума, 1975). местообитания и потреблять ресурсы. При усилении внутривидовой конкуренции происходит дифференциация вида; он занимает большую территорию, распространяясь на менее благоприятные участки ареала (рис. 6.6). Расширяется экологическая ниша. Если преобладает межвидовая конкуренция, то происходит обратное: ареал уменьшается до территории с оптимальными условиями; одновременно увеличивается специализация (однообразие) вида.
Тема 7
|
||||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 549; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 52.14.26.141 (0.015 с.) |