Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Хорологический аспект изучения экосистемыСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Выделение (разграничение) экосистем в природе часто оказывается далеко не простой задачей. Прежде всего, не существует единой точки зрения по поводу минимальной размерной единицы экосистемы. Некоторые исследователи склонны рассматривать в качестве таковой сравнительно простые, небольшие по размеру сообщества: сообщество разлагающегося ствола дерева, население верхней поверхности листа кувшинки и т. п. Действительно, принимая за основу первое из приведенных в теме 8 определение экосистемы: «...любое непрерывно меняющееся единство, включающее...», можно считать экосистемой любой биоценоз, отвечающий таким требованиям, как наличие трофических уровней, влияние на микроклимат и т. д. Но вспомним другую формулировку, в ней, в отличие от первой, заключен фактор времени: «...исторически сложившаяся система...». Видимо, «население» пня или комплекс видов-сапрофагов, живущих в лепешке навоза, правильнее рассматривать лишь как фрагменты экосистемы, существующие непродолжительное время. Их можно назвать микроэкосистемами (Р. Дажо дает им название «синузия»). Автономность микроэкосистемы относительна и существенно зависит от остальных фрагментов экосистемы. Исходя из этих рассуждений, минимальной размерной единицей экосистемы следует считать более крупные, чем микроэкосистемы, единства: луг, лес, поле, озеро и т. д. ПРИНЦИПЫ РАЗГРАНИЧЕНИЯ БИОГЕОЦЕНОЗОВ При разграничении экосистем возникает проблема выбора одного из характерных признаков, которые подразделяют на физиономические, таксономические и экологические. На основе физиономических признаков можно выделить площади с растительностью, сходной по обли- ку, если здесь сгруппированы в одинаковых пропорциях растения идентичных морфологических типов и одинакового сезонного развития. Подобный подход удобен, когда имеется один или два доминирующих вида растений, например сосна в сосняке, ель в ельнике или луговое сообщество лисохвоста и герани. Таксономические критерии базируются на численно преобладающих одиночных видах или на некоторой совокупности видов, которые называются характерными. Однако случаи, когда имеется небольшое число характерных видов, довольно редки. Чаще всего приходится иметь дело с целой группой характерных видов. В природных условиях в любую экосистему входит известное число элементов, которые могут встречаться и в других экосистемах, но к совместному существованию они оказываются способными только в одной конкретной экосистеме. Эта группа видов, или характерный набор, служит лучшим отличительным признаком экосистемы. Если характерный набор видов не проявляется, неизбежно, применение для его выявления статистических методов.
Наконец, можно осуществить разграничение экосистем по экологическим признакам, т. е. по параметрам абиотической среды. Но этот способ очень труден, так как требует огромного количества измерений. Другой источник трудностей — определение границ экосистем. В некоторых случаях переход от одной экосистемы к другой бывает резким; таковы границы между лесом и посевами, между выжженными и неповрежденными участками леса. Поскольку реально существующей единицей экосистемы является сама экосистема, а не биотическое сообщество, там, где абиотические факторы меняются резко, выделение границ экосистемы не представляет особого труда. Об изменении абиотических факторов удобно судить по видам-индикаторам, которые, будучи стенобионтными, служат критерием резкого изменения физической среды. Крупные виды являются лучшими индикаторами, чем мелкие, потому что при том же энергетическом потоке может под- дирживаться большая биомасса. Скорость оборота органического вещества у мелких организмов бывает так велика, что сдельный вид, зарегистрированный в момент исследования, может и не быть особенно полезным экологическим индикатором. В ряде случаев постепенное изменение абиотических Факторов вызывает серьезные трудности в определении границ экосистемы. Подобное явление имеет место, например, на склонах. На пологом склоне градиент абиотических факторов определяет постепенное изменение сообществ, поэтому некоторые экологи видят в них не соседствующие экосистемы, а некую непрерывность, называемую континуумом. Однако анализ биотических сообществ на территориях, достаточно ограниченных для того, чтобы единообразие сообществ было реальным, показывает относительную прерывистость и позволяет выделить на пологом склоне несколько экосистем. На крутом склоне наблюдается больший разрыв непрерывности, что облегчает определение границ экосистем.
В случае резких границ между двумя конкурирующими сообществами возникает зона напряжения, или э котон. Иными словами, экотон представляет собой зону перехода между различными сообществами, например между лесом и лугом или в море между участками с мягким и твердым грунтом. Эта пограничная зона может иметь значительную протяженность, но она всегда уже территории, прилегающих к ней экосистем. Обычно в экотонное сообщество входит значительная доля видов из перекрывающихся сообществ, а иногда также виды, характерные только для экотона. Число видов и плотность популяций некоторых из них в экотоне часто выше, чем в лежащих по обе стороны от него экосистемах. Тенденция к увеличению разнообразия и плотности живых организмов на границах сообществ известна под названием краевого эффекта. Одним из обычных и наиболее важных для человека экотонов является опушка леса. Опушку можно определить как переходное сообщество между лесным и травянистым сообщества- ми. Где бы ни жил человек, он стремится сохранить поблизости от своего жилища сообщество опушек. Так, если человек селится в лесу, он вырубает его до отдельных небольших участков, перемежающихся слугами. А поселившись на открытом месте, сажает деревья, также создавая мозаичную структуру ландшафта. Некоторые виды, обычные для леса или степи, способны выжить на опушке, созданной человеком. Другие виды, хорошо адаптировавшиеся к жизни на опушках, особенно многие виды сорняков, птиц, насекомых и млекопитающих, представлены большим числом особей. Известно, что плотность певчих птиц выше на территориях усадеб, в окрестностях поселков и других местах, которые состоят из смешанных местообитаний и, следовательно, характеризуются большей протяженностью границ по сравнению с большими однородными участками леса или полей. Таким образом, границы сообщества, находящегося в состоянии равновесия с соседними сообществами, обозначены модификацией некоторых факторов среды, которые играют решающую роль в пределах этих границ. На площади, занятой таким сообществом, особенности местообитания, напротив, претерпевают изменения, совместимые с существованием и сохранением сообщества. ИЕРАРХИЧЕСКИЙ РЯД ЭКОСИСТЕМ Установив минимальную размерную единицу экосистемы — биогеоценоз, можно построить иерархический ряд экосистем (рис. 13.1). Применяя термин «биогеоценоз» в этом смысле, мы сохраняем знак равенства между экосистемой и биогеоценозом лишь для самого низкого уровня в иерархии экосистем. Существование каждого из таких уровней определяется действием его специфического фактора. Масштаб факторов возрастает по мере перехода от низших уровней к высшим: Биосфера космический фактор Экосистемы суши и океана геологический фактор Биогеографическая область фактор эволюции Биом фактор климатического климакса (региональный климат) Ландшафт фактор рельефа Биогеоценоз фактор эдафического климата (мезоклимат).
Рис. 13.1. Иерархический ряд экосистем. Рассмотрим каждый из уровней. Процент площадей, способных поддерживать сообщества в состоянии климатического климакса, различен для разных областей. Но поскольку стратегия развития любой экосистемы состоит в достижении климатического климакса, то главными наземными экосистемами можно считать биомы. Они легко выделяются, в частности, по климатической климаксной растительности (рис. 13.2). По этому показателю можно выделить тундру, северный хвойный лес (тайгу), лиственный лес умеренной зоны, степь, пустыню, тропический дождевой лес и т. д.
В разных биогеографических областях сообщества сильно различаются по видовому составу. Каждый вид образуется в одном, определенном месте земного шара, а затем расселяется, останавливаясь перед естественными преградами, такими как морские проливы, горные цепи и т. д. Однако всюду, где независимо от географического положения физическая среда одинакова, развиваются сходные экосистемы. Эквивалентные экологические ниши оказываются занятыми теми биологическими
Рис. 13.2. Основные биомы европейской части СССР (из Сукачева, 1934, с изменениями). Там, где кривая осадков пересекает восходящую линию испаряемости, расположена граница между гумидным (слева) и аридным (справа) климатом. группами, которые имеются в фауне и флоре данной, области. Так, степной биом развивается во всех областях со степным климатом, но виды злаков и травоядных животных могут быть различными. Организмы, занимающие одинаковые или сходные экологические ниши в разных географических областях, называются экологическими эквивалентами. Кенгуру в Австралии — экологический эквивалент бизона и вилорогой антилопы в Северной Америке. Виды злаковых трав, которыми питаются травоядные, внешне очень сходны по всему земному шару, хотя конкретные виды, а также роды и даже семейства могут быть строго приурочены к данному материку или к определенной биогеографической области в его пределах. Огромные различия в физико-химических свойствах между наземными и водными средами создают в них совершенно разные условия жизни. Своеобразие экосистем океана в отличие от экосистем суши определя- ется в первую очередь абиотическими факторами, а также рядом особенностей общего характера: 1) море занимает 70% поверхности Земли; 2) глубина моря огромна и жизнь обнаруживается в пределах всей этой толщи. В океане, по-видимому, не существует абиотических зон, однако воды вблизи материков и островов населены значительно обильнее; 3) море в отличие от суши и пресных вод непрерывно. Все океаны связаны друг с другом. Основными барьерами для свободного передвижения морских животных служат температура, соленость и глубина;
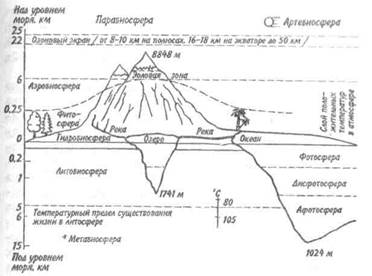
4) в море происходит постоянная циркуляция воды. Разница в температуре воздуха на полюсах и у экватора порождает сильные ветры, не меняющие своего направления на протяжении всего года (пассаты). В результате совместного действия этих ветров и вращения Земли образуются определенные течения. Каждый из иерархических уровней экосистем следует рассматривать не только как систему, объединяющую подразделения низшего порядка, но и как самостоятельную экосистему, обладающую всеми присущими ей свойствами. В зависимости от размера биотопа можно выделить экосистемы различного порядка. Так, биогеоценоз — это экосистема низшего порядка, а биосфера—экосистема высшего, или первого, порядка. Если рассматривать сравнительно ограниченный биотоп с более или менее однообразными абиотическими факторами, например луг, лес или участок морского дна, то количество видов позвоночных, входящих в состав биоценоза, определяется небольшой величиной —50-200 видов. Для более крупных биотопов, например морей умеренной зоны, количество видов позвоночных животных составляет 1-2, а для теплых морей оно возрастает до 5-8 тыс. БИОСФЕРА Понятие биосферы вошло в науку случайно. Более 100 лет назад, в 1875 г., австрийский геолог Эдуард Зюсс, говоря о различных оболочках земного шара, впервые упот- ребил этот термин в последней, наиболее общей главе своей книги о происхождении Альп. Однако это упоминание не сыграло сколько-нибудь заметной роли в развитии научной мысли. В 1926 г. были опубликованы две лекции русского минералога В. И. Вернадского, в которых им, спустя 50 лет после работ Зюсса, формулировались основные положения концепции биосферы, которую мы принимаем и сейчас. Под биосферой Вернадский понимал те слои земной коры, которые подвергались в течение всей геологической истории влиянию живых организмов. В последние годы многие ученые (Дж. Хатчинсон и др.) сужают представление о биосфере, рассматривая ее как ту часть поверхности Земли, которая в настоящее время находится под влиянием деятельности организмов. Многие научные термины в разных случаях применяются то в более широком, то в более узком понимании. Что же характерно для биосферы — этой особой оболочки земного шара? Во-первых, в биосфере весьма значительно количество жидкой воды. Во-вторых, на нее падает мощный поток энергии Солнца. В-третьих, для биосферы характерны поверхности раздела между веществами, находящимися в жидком, твердом и газообразном состояниях. Поскольку источником энергии на Земле является Солнце, то все живые организмы распределены в верхних слоях двух земных оболочек: литосферы и гидросферы (рис. 13.3). Чем лучше та или иная земная оболочка пропускает солнечные лучи, тем на большую глубину она заселена живыми организмами. Однако биосфера не кончается там, куда не доходит свет. Благодаря силе тяжести поток энергии распространяется еще дальше: из освещенных слоев в глубину моря непрестанно падают комочки экскрементов, мертвые и живые организмы.
В литосферу живые организмы проникают на ничтожную глубину. Основная их масса сосредоточена в верхнем слое почвы мощностью в несколько десятков сантиметров, и редко кто проникает на несколько метров или десятков метров вглубь (корни растений, дождевые
Рис. 13.3. Строение биосферы. черви). По трещинам земной коры, колодцам, шахтам и буровым скважинам животные и бактерии могут опускаться на гораздо большую глубину—до 2,5-3 км. Нефть, часто залегающая глубоко от поверхности земли, имеет своеобразную бактериальную флору. Проникновение зеленых растений в глубь литосферы невозможно из-за отсутствия света. Животные не находят там питания. Механические свойства горных пород, слагающих литосферу, также препятствуют распространению в них жизни. Наконец, с продвижением в недра Земли температура возрастает и на глубине 3 км достигает 100 °С. Значит, на глубине более 3 км от земной поверхности живые организмы существовать не могут. С поверхности литосферы живые организмы проникают в нижние слои атмосферы — на высоту от нескольких сантиметров до нескольких метров. А растения возносят свои зеленые кроны иногда на несколько десятков метров. На несколько сотен метров в атмосферу проникают насекомые, летучие мыши и птицы. Восходя- щие токи воздуха могут поднимать на несколько километров покоящиеся стадии (споры, цисты, семена) животных и растений. Однако организмы, проводящие всю свою жизнь в воздухе, т. е. связанные с ним как с основной средой обитания, не известцы. Гидросфера в отличие от атмосферы и литосферы заполнена жизнью по всей своей толще. Повсюду, куда проникали орудия сбора, исследователи находили живые организмы. Из этого мы можем заключить, что жидкая вода является более важным лимитирующим фактором в расселении организмов, чем свет. Так, самые жаркие пустыни формально находятся вне биосферы. Однако фактически они могут считаться парабиосферными (околобиосферными), так как живые организмы там все же есть. Например, в пустынях Намиб и Калахари под слоем сухого песка встречаются насекомые (жуки-чернотелки), существующие за счет приносимых ветром сухих пылевидных остатков растений; питаясь ими, насекомые получают метаболическую воду. Протяженность биосферы ввысь ограничена в основном недостатком жидкой воды и низким парциальным давлением углекислого газа. В горах хлорофиллсодержащие растения, видимо, не могут жить на высоте более 6200 м (Гималаи). На еще больших высотах встречаются некоторые животные, например пауки. Они питаются ногохвостками, а те, в свою очередь, довольствуются зернами пыльцы, заносимыми сюда ветром. Высокогорную область биосферы называют эоловой зоной. Если лимитирующими факторами биосферы являются жидкая вода и солнечный свет, то оптимум жизни приходится на поверхность раздела сред. Исследования фотосинтеза показали, что часто наибольший выход органических веществ дают растения, способные использовать все три фазы: твердую, жидкую и газообразную. Примером может служить тростник обыкновенный, Phragmites communis. Всасывание воды для него облегчается постоянным давлением жидкости на донные осадки. Необходимый для существования углекислый газ тростник получает из газообразной среды, в которой скорость проникнове- ния газа через поглощающие поверхности наиболее высока; кислород тоже легче получать из воздуха, чем из воды; наконец, все остальные элементы легче извлекать из раствора в капиллярной воде осадка. Предпринимались многочисленные попытки оценить первичную продукцию биосферы. Обширные пространства Земли попадают в категорию низкопродуктивных из-за таких лимитирующих факторов, как вода (в пустынях) или питательные вещества (в открытом море). Хотя площадь суши составляет всего около 1/4 общей площади планеты, суша превосходит океаны по своей продуктивности, так как большая часть океанских вод в основном «пустынна» (таблица). В Мировом океане значения первичной продукции в разных районах существенно различаются. Наиболее продуктивны коралловые рифы, не уступающие даже тропическим лесам. Продуктивность открытых океанических зон ниже продуктивности зон апвеллинга и прибрежных районов и близка к таковой для тундры. Анализ оценок средних величин для больших площадей показывает, что продуктивность колеблется в пределах двух порядков— от 200 до 20 000 ккал на 1 м2 в год, а общая валовая продукция Земли имеет величину порядка 1018 ккал в год. Чтобы биосфера продолжала существовать, не должен прекращаться круговорот основных химических элементов. Скорость этих процессов может быть различной. Органическое вещество, которое ведет свое происхождение от атмосферного диоксида углерода, завершает цикл формирования за время, исчисляемое десятилетиями. Круговорот кальция совершается значительно медленнее. В форме бикарбоната Са(НС03)2 кальций смывается в реки из континентальных пород, осаждаясь затем в виде карбоната СаС03 в водах открытого океана главным образом в форме тончайших раковин фораминифер. Дно океана медленно перемещается к поясам горообразования на окраинах материков, и кальций возвращается на сушу. Круговорот завершается, по-видимому, за несколько сотен миллионов лет. Фосфор, как мы знаем, по характеру своего круговорота схож с каль- цием, а азот имеет больше сходства с углеродом, хотя его содержание в атмосфере гораздо выше. В разных участках биосферы развитие жизни лимитируется разными веществами. Можно сказать, что в пустыне жизнь ограничена недостаточным количеством водорода и кислорода в форме воды. В открытом океане лимитирующим фактором часто служит железо, обычно присутствующее в форме труднодоступной для организмов гидроокиси. В иных средах, например в почвах влажных районов, в озерах, окраинных морях, лимитирующим фактором чаще всего является фосфор.
|
|||||||||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 626; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.222.127.152 (0.014 с.) |