
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Внутренняя организация биотического сообществаСодержание книги
Поиск на нашем сайте Облик биотического сообщества определяется не только разнообразием видов и другими показателями, которые отражают связи между видами, входящими в состав биотического сообщества. Функционирование сообщества и его стабильность зависят также от популяционных связей, от распределения организмов в пространстве и характера их взаимодействия с внешней средой. Все это составляет понятие внутренней организации сообщества. О ней можно судить на основании следующих параметров. Стратификация (вертикальная ярусность). В лесу два основных яруса — автотрофный и гетеротрофный — часто подразделяются на дополнительные. Так, растительность может быть представлена травами, кустарниками, древесными ярусами; почва также отчетливо подразделяется на горизонты. Стратификация выражена не только в лесных и водных экосистемах. Даже в такой, казалось бы, «одномерной» экосистеме, как луг, можно выделить несколько ярусов: почву, где проводят всю жизнь дождевые черви, личинки жуков и другие животные; поверхность почвы, к которой приурочены муравьи, жужелицы; слой мха, где обитают первичнобескрылые насекомые и панцирные клещи; стебли и листья, с которыми связаны кузнечики, тли и другие животные; цвет- ки, на которых проводят много времени цветочные пауки, шмели и др. Зональность (горизонтальные подразделения). Растения и животные распределены не равномерно по всей экосистеме, а пятнами, в которых плотность может быть максимальной или, наоборот, сходить на нет. Активность (периодичность). Периодичность сообщества является результатом синхронной активности в течение дня и ночи целых групп организмов. Некоторые из них активны только ночью, другие — днем, а третьи — только в сумерки. Поразительным примером суточной периодичности в водоемах служит вертикальная «миграция» зоопланктона в морях и озерах: веслоногие и ветвистоусые рачки ночью движутся вверх, к поверхности воды, а днем — вниз. Для всех сообществ характерна также сезонная периодичность, что нередко приводит почти к полному изменению структуры сообщества в течение года. Хотя принято различать четыре времени года, деление года на шесть времен больше соответствует периодичности сообществ средних широт: 1) зимняя спячка (зима), 2) начало пробуждения (ранняя весна), 3) пробуждение и высокая активность (поздняя весна), 4) максимальная активность (раннее лето), 5) окончание активности (позднее лето), 6) подготовка к зимней спячке (осень). Размножение (связи потомков с родителями, половое, бесполое размножение, партеногенез и пр.). Пищевые связи (сетчатая структура пищевых связей). Групповые отношения (стаи, стада). Совместная деятельность (конкуренция, комменсализм, мутуализм и т. д.).- Стохастические связи (случайные воздействия). Подробнее последние пять параметров организации сообщества обсуждались в соответствующих разделах рассмотренных выше тем.
Те м а 12 РАЗВИТИЕ И ЭВОЛЮЦИЯ ЭКОСИСТЕМЫ Одно из основных свойств экосистем - их динамизм. Наблюдение над полем, заброшенным на несколько лет, показывает, что его последовательно завоевывают сначала многолетние травы, затем кустарники и, наконец, древесная растительность. Изменение экосистем может происходить под воздействием разных причин. В зависимости от вектора действующих сил различают аллогенные и автогенные изменения. Аллогенные изменения обусловлены влиянием геохимических сил, действующих на экосистему извне. В качестве таковых могут выступать климатические и геологические факторы. Влияние климатических факторов наглядно видно на примере тех изменений, которые имели место в Европе в четвертичное время в течение ледникового и межледникового периодов. При максимальном развитии ледника Средняя Европа представляла собой тундру с карликовыми ивами, дриадами и камнеломками, а вся флора умеренного климата была вытеснена на крайний юг. Ледниковая фауна состояла из мамонтов, волосатых носорогов, мускусных овцебыков и мелких грызунов, таких как лемминги. Потепление, наступавшее в межледниковые периоды, позволяло теплолюбивым растениям, в частности винограду, возвращаться в районы к северу от Альп, а теплолюбивой фауне, в том числе древнему слону и гиппопотаму, обосноваться в Европе. Такие геологические явления, как эрозия, образование осадочных пород, горообразование и вулканизм, могут настолько изменять физическую среду, что вызывают значительные сдвиги в экосистемах. В отличие от аллогенных автогенные изменения обусловлены воздействием процессов, протекающих внутри экосистемы. В большинстве случаев, однако, трудно разграничить процессы, находящиеся под влиянием внешних и внутренних факторов. Например, эвтрофикация озер
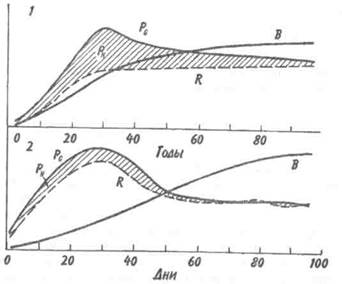
происходит под действием населяющих их сообществ, толчком к изменению которых служит поступление в озеро питательных веществ извне, с водосбора. Тем не менее, степень участия сообщества в преобразовании экосистемы, как правило, устанавливается без особого труда и, кроме того, автогенные изменения характеризуются рядом признаков. ЭКОЛОГИЧЕСКАЯ СУКЦЕССИЯ Автогенные изменения называют развитием экосистемы, или экологической сукцессией. При определении экологической сукцессии следует учитывать три момента: 1. Сукцессия происходит под действием сообщества, т. е. биотического компонента экосистемы. Сообщество, в свою очередь, вызывает изменения в физической среде, которая определяет характер сукцессии, ее скорость и устанавливает пределы, до которых может дойти развитие. 2. Сукцессия — это упорядоченное развитие экосистемы, связанное с изменением видовой структуры и протекающих в сообществе процессов. Сукцессия определенным образом направлена и, следовательно, предсказуема. 3. Кульминацией сукцессии является возникновение стабилизированной экосистемы, в которой на единицу потока энергии приходится максимальная биомасса и максимальное количество межвидовых взаимодействий. Скорость изменений и время, необходимое для достижения стабилизированного состояния, варьируют в разных экосистемах и для разных признаков экосистемы. Кривая изменения общей биомассы обычно имеет выпуклую форму, тогда как кривая изменения валовой первичной продукции — вогнутую (рис. 12.1). Как в лесной экосистеме, так и в условиях лабораторного микрокосма чистая первичная продукция возрастает на ранних стадиях развития сообщества, но убывает — на поздних. Оценка изменений этих и других параметров приведена в табл. 12.1.
Рис. 12.1. Энергетика развития экосистемы леса (/) и лабораторного микрокосма (2) (по: Одум, 1975). Остальные обозначения см. на рис. 9.4. В — общая биомасса. На ранних стадиях экологической сукцессии валовая первичная продуктивность превосходит затраты на дыхание сообщества, так что Р/R >1. Для особого случая, т. е. для случая сильного органического загрязнения среды (например, в сточных водах), затраты на дыхание могут превалировать над валовой первичной продуктивностью. Если сукцессия начинается с подобного состояния, то говорят о гетеротрофной сукцессии в отличие от обычной, авто-трофной сукцессии. Однако в любом случае по мере развития экосистемы отношение P/R стремится к 1. Графически это можно изобразить в виде прямой, составляющей угол 45 ° с осями координат (рис. 12.2), несмотря на то, что в разных экосистемах, в зависимости от физических факторов среды, абсолютные значения продуктивности и, следовательно, затрат на дыхание сообщества могут существенно различаться. Пока Р больше R, в системе будут накапливаться органическое вещество и биомасса, в результате чего отно-
Рис. 12.2. Положение сообществ различных типов в классификации, основанной на метаболизме (по: Одум, 1975). Направления автотрофной и гетеротрофной сукцессии указаны стрелками. Сообщества, расположенные вдоль диагонали, потребляют за год в среднем примерно столько, сколько они создали, и их можно считать метаболически климаксными. шение Р/В будет снижаться, а отношения В/Р, B/R или В/Е (где Е= Р+ R) соответственно увеличиваться. Иными словами, биомасса, поддерживаемая доступным потоком энергии, достигает максимума в зрелых экосистемах (табл. 12.1, признак 3). Как следствие этого, чистая продукция сообщества — урожай за годовой цикл — велика на ранних стадиях развития экосистемы и мала или равна нулю в зрелом ее состоянии (признак 4). По мере развития экосистемы следует ожидать тонких изменений в структуре пищевых цепей (признак 5). При ненарушаемом течении сукцессии имеется достаточно времени для развития более тесных связей и взаимных адаптации между животными и растениями, что ведет к появлению множества механизмов, уменьшающих выедание растений: образуются плохо перевариваемые опорные ткани, увеличивается давление хищников на фитофагов и т. д. Листва опадает, и переработка органического вещества переходит из зеленого пояса в коричневый.
В развивающихся системах мало число видов и мала их выравненность, не развиты гетеротипические реакции, а в зрелых - число видов и их выравненность велики, гетеротипические реакции развиты (признаки 9 -11). Особенно примечательны в этом отношении тесные связи между неродственными видами, например между кораллами (кишечнополостные) и зооксантеллами (одноклеточные жгутиковые) или между микоризой и деревьями. Как следствие усиления гетеротипических реакций происходит специализация по нишам и усиление стратификации (признаки 12, 13). Рассмотрим подробнее признаки 14 и 15. На начальной стадии сукцессии организмы обычно бывают мелкими, для них характерны простые жизненные циклы и высокие скорости размножения. Изменения размеров особей происходят вследствие перемещения биогенных веществ из неорганической в органическую фазу. Малые размеры дают преимущество, особенно автотрофам, в среде, богатой минеральными веществами, поскольку возрастает отношение площади поверхности тела к его объему. Однако, по мере развития экосистемы, неорганические биогенные вещества становятся все более связанными в биомассе, так что преимущество переходит к организмам с большим объемом, т. е. к более крупным — либо к более крупным особям данного вида, либо к видам с более крупными особями, либо к тем и другим, так или иначе обладающим большими возможностями для накопления веществ и более сложными жизненными циклами. Характер роста популяции меняется от экспоненциального к логистическому, так как возрастает сопротивление среды. Известно, что в ограничении роста популяции, когда увеличение ее численности происходит по логистическому закону, решающее значение имеет отрицательная обратная связь. Во многих случаях биотическая регуляция выедания растительности (признак 5), плотности популяции (признак 16) и круговорота веществ (признак 6) служит основным механизмом действия отрицательной обратной связи. В результате расширения поля деятельности для отрицательной обратной связи увеличивается стабильность экосистемы (признак 17), снижается ее энтропия (признак 18) и в конечном итоге повышается количество содержащейся в ней информации (признак 19). При загрязнении или иной форме стресса вектор изменения признаков экосистемы становится противоположным. В этом легко убедиться, анализируя изменения, которые сопровождают значительное загрязнение нефтепродуктами малых рек Западной Сибири. Это обеднение видового состава во всех звеньях пищевых цепей (22 вида в фитопланктоне вместо 31; 4 вида, высших растений вместо 28; 1 вид ветвистоусых ракообразных в зоопланктоне вместо 5; 20 видов в бентосе вместо 45 и т. д.), концентрация доминирования в немногих видах, устойчивых к загрязнению, уменьшение стратификации (слоистости) сообщества, т. е. равномерности распределения организмов по микробиотопам, уменьшение суммарной биомассы (либо в результате снижения видового разнообразия, либо в результате того, что особи видов, устойчивых к нефтяному загрязнению, имеют меньший индивидуальный вес), сужение кормовой базы рыб вследствие угнетения зоопланктона, животных-—обитателей зарослей высших водных растений, и концентрации биомассы в мезобентосе. Таким образом, знание изменений, которые происходят в процессе экологической сукцессии, во многих случаях открывает возможность для прогноза неблагоприятных последствий антропогенного воздействия на экосистемы. Основной результат деятельности сообщества состоит в усилении гетеротипических реакций, сохранении минеральных веществ, повышении стабильности и увеличении информации. Этот результат называется всеобщим гомеостазом. Стратегия экологической сукцессии направлена на достижение такой обширной и разнообразной органической структуры, какая только возможна в границах, установленных доступным потоком энергии и физическими условиями существования.
|
||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 480; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.170 (0.009 с.) |




 Важной тенденцией в развитии экосистемы является стремление к замыканию биогеохимических круговоротов основных элементов, таких как азот, фосфор и кальций. Зрелые системы по сравнению с развивающимися обладают большей способностью захватывать вещества и сохранять их в обменном фонде. Они теряют лишь очень небольшие количества веществ по сравнению с незрелыми или нарушенными системами. Отсюда следуют изменения, отмеченные в табл. 12.1 (признаки 6-8).
Важной тенденцией в развитии экосистемы является стремление к замыканию биогеохимических круговоротов основных элементов, таких как азот, фосфор и кальций. Зрелые системы по сравнению с развивающимися обладают большей способностью захватывать вещества и сохранять их в обменном фонде. Они теряют лишь очень небольшие количества веществ по сравнению с незрелыми или нарушенными системами. Отсюда следуют изменения, отмеченные в табл. 12.1 (признаки 6-8).


