

Мы поможем в написании ваших работ!
ЗНАЕТЕ ЛИ ВЫ?
|
Участие почек в поддержании кислотно-основного равновесия крови.
Содержание книги
- Рефлекторная функция спинного мозга
- Проводниковая функция спинного мозга
- Спинальные механизмы регуляции мышечного тонуса и фазных движений.
- Клинически важные спинальные рефлексы
- Рефлекторная функция продолговатого мозга.
- Рефлекторная функция продолговатого мозга. Бульбарные механизмы поддержания позы человека.
- Структурно-функциональная организация и физиологические функции среднего мозга и моста (сенсорные, проводниковые, моторные, вегетативные, интегративные, рефлекторные).
- Современные представления о влиянии РФ.
- Структурно-функциональная организация лимбической системы. Ее роль в формировании мотиваций, эмоций, организации памяти. Участие лимбических структур в интегративной деятельности цнс.
- Проводниковый отдел и принципы его построения.
- Основные функции анализаторов
- Основные формы нарушения цветового восприятия.
- Механизм работы вестибулярного анализатора
- Внимание, его нейрофизиологические механизмы. Роль внимания в процессах запоминания и обучения.
- Нейроструктурные предпосылки мышления
- Механизмы долговременной памяти
- Функциональная асимметрия полушарий головного мозга и ее роль в реализации психических функций
- Механизмы возрастания минутного объёма дыхания при физической нагрузке. Причины развития дыхательного алкалоза при тяжёлой мышечной работе.
- Чем представлен дыхательный аппарат у человека? Респираторные и нереспираторные функции воздухопроводящих путей и легких.
- Перечислите этапы биомеханики вдоха. Назовите виды давления в грудной полости и их роль в биомеханике вдоха и выдоха?
- Специфические регуляторы дыхания: опишите зависимость минутного объёма дыхания от рН ликвора, рО2 в крови. Опыт Фредерика.
- Работа дыхательных мышц и её зависимость от сопротивления дыханию. Виды сопротивления. Что такое предел дыхания?
- Механизм спонтанного дыхания
- Периоды рефрактерности сердца
- Резус-фактор, физиологическая роль. Физиологические основы переливания крови. Основные правила переливания крови. Гемотрансфузионный шок.
- ФУС, обеспечивающая поддержание постоянства уровня питательных веществ в крови.
- Пищеварение в полости рта. Состав и пищеварительные свойства слюны. Регуляция секреторной функции слюнных желез.
- Пищеварение в желудке. Состав и пищеварительные свойства желудочного сока. Фазы желудочной секреции. Механизмы регуляции желудочной секреции.
- Состав и пищеварительное действие поджелудочного сока. Регуляция панкреатической секреции.
- Моторная функция желудка, тонкого и толстого кишечника. Физиологические особенности и значение .
- Механизмы всасывания. Виды всасывания. Виды транспорта веществ через мембрану. Особенности всасывания углеводов, белков, жиров.
- Участие почек в поддержании кислотно-основного равновесия крови.
- Строение нефрона. Кровообращение в почке, его особенности.
- Реабсорбция в почечных канальцах. Виды реабсорбции. Механизмы реабсорбции. Пороговые вещества. Регуляция реабсорбции.
- Температурное «ядро» и «оболочка». Методы измерения температуры тела. Количество тепловой энергии, вырабатываемой в сутки в организме теплокровного животного, механизмы её распределения в организме.
- Способы отдачи тепла (теплопроведение, конвекция, излучение, испарение). Внутренний и наружный потоки энергии. Регуляция теплоотдачи.
- Терморегуляция. Понятие о гипотермии и гипертермии.
- Механизмы адаптации к теплу и холоду.
- Учёт расхода энергии в организме. Прямая и непрямая калориметрия.
- Методы прямой и непрямой калориметрии.
- Основной обмен, величина и факторы, его определяющие. Правило поверхности тела.
- Рабочий обмен. Группы людей по энергозатратам. Специфически-динамическое действие питательных веществ.
- Основные системы питания, теории рационального питания. Значение белков, жиров, углеводов.
- Физиологические нормы питания. Понятие о белковом минимуме и белковом оптимуме. Нормы углеводов и жиров для человека. Соотношение питательных веществ в пищевом рационе.
- Методы изучения желез внутренней секреции
- Нейросекреты гипоталамуса. Либерины и статины.
- Тиреоидные гормоны и их эффекты.
- Роль гормонов щитовидной и паращитовидной желез в регуляции уровня кальция.
- Инкреаторная функция поджелудочной железы.
- Гормоны мозгового вещества надпочечников и их роль.
Почки играют важную роль в поддержании постоянства концентрации Н+ в крови, экскретируя кислые продукты обмена.
Механизм подкисления мочи основан на секреции клетками канальцев ионов водорода.
Причины, определяющие, будет ли моча кислая:
1) Постоянно фильтруется большое количество бикарбонатов в первичную мочу. Это есть механизм удаления оснований из крови.
2) В то же самое время в просвет канальцев секретируется большое количество Н+ при помощи эпителиоцитов. Это есть механизм потери кислот из внеклеточной жидкости.
Секреция Н+. Клетки проксимальных и дистальных канальцев, вроде клеток желудочных желез секретируют Н+. Наиболее важное значение в секреции Н+ в проксимальных канальцах имеет Н+/Nа+ обмен (вторично-активный транспорт). Благодаря Nа+-К+-АТФазе внутриклеточная концентрация ионов, Nа+ поддерживается на низком уровне. Это заставляет входить Nа+ из мочи в эпителиоциты по градиенту концентрации. В свою очередь, вход Nа+ сопряжен с выходом Н+ в мочу. В эпителиоцитах Н+ образуется при распаде Н2СО3 на НСО3- и Н+. Следовательно, в обмен на один атом водорода, входящий в клетку, из нее выходит Nа+ и НСО3 в межклеточную жидкость.
В дистальных канальцах и собирательных трубочках секреция водорода в мочу обусловлена работой водородного насоса, на работу которого затрачивается АТФ. Активируя этот насос альдостерон усиливает секрецию Н+ в дистальных канальцах.
Регуляция секреции H+ концентрацией H+во внеклеточной жидкости.
Скорость секреции Н+ в каналъцах существенно изменяется в ответ даже на небольшое изменение Н+ во внеклеточной жидкости.
Следовательно, когда концентрация Н+ очень высокая (рН меньше, чем 4,5) его секреция может увеличиваться в 7 раз и наоборот.
Это достигается следующим образом:
1. При ацидозе отношение СО2 /НСО3- увеличивается по сравнению с нормой. Точно такое же взаимоотношение между СО2 и НСО3- имеет место в эпителиоците, секретирующем Н+ в просвет канальца и, следовательно, секреция Н+ возрастает. При алкалозе преобладают НСО3- анионы и секреция Н+ уменьшается.
2. В норме скорость экскреции ионов водорода равна 3,5 мМ/мин, но эта величина может уменьшаться или увеличиваться в зависимости от концентрации Н+ во внеклеточной жидкости.
Взаимодействие НСО3- и Н+ в просвете почечных канальцев. Реабсорбция ионов бикарбоната. Эпителий почечных канальцев мало проницаем для НСО3-. Однако, он эффективно реабсорбируется.
Реабсорбция НСО3- начинается с взаимодействия НСО3-, фильтрующегося в клубочках и Н+, секретируемого эпителиоцитами почечных канальцев. В результате образуется Н2СО3, которая диссоциирует на СО2, и Н2О. СО2 затем по градиенту концентрации проходит через люминальную мембрану эпителиоцита, а вода остается в просвете канальца. В эпителиоците СО2 взаимодействует с Н2О и вновь образуется Н2СО3, которая диссоциирует на Н+ и НСО3. Следовательно, в результате этой реакции образуется вновь Н+, который будет секретироваться в просвет канальца, а НСО3 диффундирует через базалъную мембрану эпителиоцита во внеклеточную жидкость.
Титрование ионов бикарбоната ионами водорода. В нормальных условиях Н+ секретируется со скоростью 3,5 мМ/мин, а скорость фильтрации НСО3- анионов в клубочках равна 3,46 мМ/мин. Следовательно, количество этих двух ионов, поступающих в просвет каналъца почти равны, поэтому они взаимодействуют и нейтрализуют друг друга.
Однако, количество Н+ и НСО3- не совсем эквивалентно, поэтому небольшой избыток ионов Н+ остается в просвете канальцев и попадают в мочу подкисляя ее.
Если фильтруется большое количество НСО3-, то часть его идет на нейтрализацию водородных ионов, а часть поступает во вторичную мочу, делая ее щелочной.
Таким образом, основным механизмом, посредством которого почка коррегирует ацидоз или алкалоз является неполное титрование водородных ионов ионами бикарбоната.
Механизмы связывания большого количества ионов водорода, поступающих в каналъцевую мочу. После того как большая часть водородных ионов секретируется в просвет какальца лишь небольшая их часть попадает в мочу. Причины этого следующие:
- максимальная концентрация ионов Н+, которую может создать канальцевая система секреции Н+ равна 10-4,5 моль, т.е. рН = 4,5. Следовательно, чтобы перенести избыток, Н+ в мочу ионы водорода должны поступать в нее не в свободном виде. Это достигается тем, что Н+ в просвете канальца взаимодействует с внутриканальцевыми буферными системами и затем в связанном виде выводится.
- В канальцевой жидкости имеются две важные буферные системы: а) аммонийный буфер и б) фосфатный буфер. Кроме того имеются многочисленные слабые буферные системы: уратная, цитратная и др. Их значение невелико.
Транспорт избытка ионов водорода в мочу при помощи фосфатного буфера. Фосфатный буфер состоит из НРО42- и Н2РО4. Оба эти компонента буферной значительно концентрируются в канальцевой жидкости. Это связано с тем, что 1) фосфаты реабсорбируются незначительно, а вода в большом количестве. Поэтому относительно слабый фосфатный буфер крови в моче приобретает важное значение; 2) возрастание значения фосфатного буфера в моче связано с тем, что в дистальном сегменте моча имеет рН = 6,0, а именно при этой рН эффективность данного буфера велика.
Отметим, что когда ион водорода связывается с гидрофосфатом, в канальце, то один ион НСО-3 поступает в кровь.
Транспорт избытка ионов водорода в мочу при помощи аммонийного буфера. Эпителиальные клетки всех канальцев за исключением тонкого сегмента петли Генли, постоянно сентизирует аммоний и он диффундирует в просвет канальца. После того, как он попадает в мочу, он взаимодействует с ионами Сl- и другими анионами, находящимися в просвете канальца.
Окончательным эффектом этих реакций снова является увеличение концентрации, бикарбонатов во внеклеточной жидкости. Значение аммонийного буфера:
- Как только аммиак взаимодействует с Н+ его концентрация в канальцевой моче уменьшается и он диффундирует из эпителиоцита в канальцевую жидкость. Следовательно, скорость диффузии аммония в канальцевую жидкость зависит от количества водородных ионов транспортируемых в каналец.
- Большинство отрицательных ионов в канальцевой моче составляют ионы Сl-. Лишь небольшое количество может взаимодействовать с Сl- с образованием соляной кислоты, что должно было бы привести к значительному снижению рН. Однако, когда Н+ взаимодействует с NH3, образуется NH4, который взаимодействует с С1- с образованием очень слабой кислоты.
· Участие почек в поддержании азотистого баланса
В процессе метаболизма белков и нуклеиновых кислот образуются различные продукты азотистого обмена (у человека — мочевина, мочевая кислота, креатинин и др.). Катаболизм пуриновых оснований в организме человека останавливается на уровне образования мочевой кислоты, в клетках некоторых животных имеются ферменты, обеспечивающие распад пуриновых оснований до СО2 и аммиака. Мочевая кислота в почке человека фильтруется в клубочках, затем реабсорбируется в канальцах, часть мочевой кислоты секретируется клетками в просвет нефрона. Обычно экскретируемая фракция мочевой кислоты довольно низкая (9,8 %), что указывает на реабсорбцию значительного количества мочевой кислоты в канальцах. Интерес к изучению механизмов транспорта мочевой кислоты в почечных канальцах обусловлен резко возросшей частотой заболевания подагрой, при которой нарушен обмен мочевой кислоты.
Образующийся в течение суток креатинин, источником которого служит креатинфосфорная кислота, выделяется почками. Его суточная экскреция зависит не столько от потребления мяса с пищей, сколько от массы мышц тела. Креатинин, как и мочевина, свободно фильтруется в почечных клубочках, с мочой выводится весь профильтровавшийся креатинин, в то время как мочевина частично реабсорбируется в канальцах.
Помимо перечисленных, имеется много разнообразных веществ, постоянно удаляемых почкой из крови. О том, какие вещества удаляет или разрушает почка, можно судить при изучении состава крови у людей с удаленными почками. В их крови, помимо мочевины, креатинина, мочевой кислоты, накапливаются гормоны (глюкагон, паратгормон, гастрин), ферменты (рибонуклеаза, ренин), производные индола, глюкуроновая кислота и др.
· Участие почек в поддержании осмотического давления крови

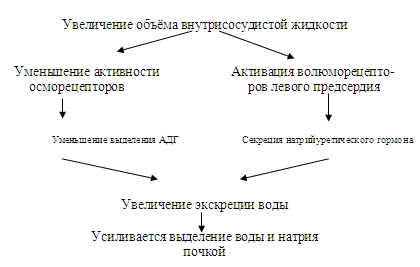
Помимо информации от осморецепторов и натрийрецепторов уровень секреции АДГ зависит от активности волюморецепторов, реагирующих на изменение объёма внутрисосудистой и внеклеточной жидкости (рис.48). К этим рецепторам относятся:
· рецепторы, реагирующие на растяжение сосудистой стенки,
· рецепторы левого предсердия, от них возбуждение передается в ЦНС по блуждающему нерву.
При увеличении кровенаполнения левого предсердия активируются волюморецепторы и угнетается секреция АДГ и усиливается мочеотделение.
В предсердии вырабатывается натрийуретический пептид, увеличивающий выделение ионов Na+ почкой.
Стимуляция волюморецепторов рефлекторно изменяет характер импульсации, поступающей по иннервирующим почку симпатическим нервом и снижает реабсорбцию Na+ и Н2О в них.
Растяжение афферентной артериолы клубочка, увеличившимся объемом крови, сопровождается изменением активности ЮГА. Уменьшается секреция ренина, а следовательно, концентрация ангиотензина II и альдостерона. Всё это увеличивает выделение Nа+почкой.
|




