Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Окклюзия развития пачечных разрядов в поле са1 в ответ на кратковременные увеличения внеклеточной концентрации калия в моделях эпилептогенеза in VIvoСодержание книги
Поиск на нашем сайте
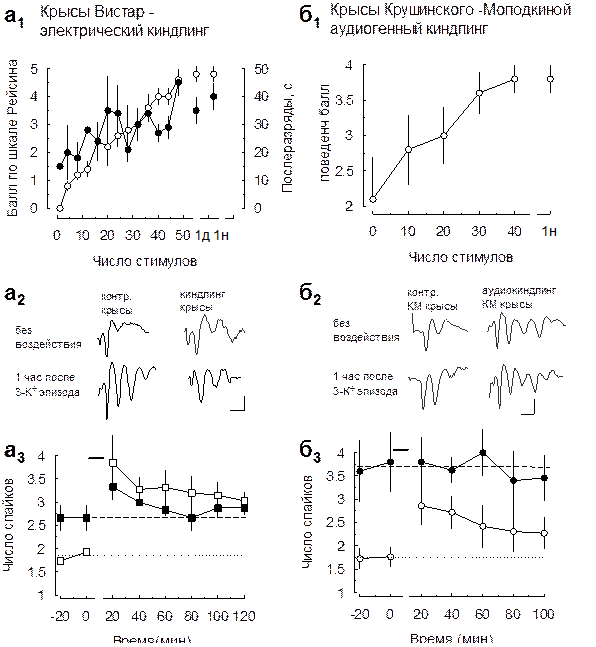
Из полученных данных осталось не ясно, является ли множественный разряд в пирамидных клетках поля СА1 эпилептиформным. Как отмечалось ранее, пачечная активность не всегда представляет собой патологический процесс, а может принимать участие в генерации мозгом нормальных ритмов (D'Angelo et al. 2001; Twery et al. 1992). Чтобы ответить на этот вопрос, мы исследовали особенности развития пачечной активности вызываемой повышением К+ в срезах гиппокампа крыс после электрического и аудиогенного киндлинга in vivo. Электрический киндлинг in vivo Проводимая через сутки (в течение 8 суток) хроническая электрическая стимуляция (частота стимуляции 50 Гц в течение 10 сек., 12 стимулов/день с интервалом 30 мин.) гиппокампа in vivo приводила к прогрессирующему увеличению длительности послеразрядов и соответствующему изменению паттерна двигательной судорожной активности в соответствии с критериями Рэйсина (Рис. 3.6.2а1). 48 таких стимуляций приводили к увеличению длительности послеразрядов свыше 30 сек. и достижению животными 4-5 стадии киндлинга. Для исследований брали срезы гиппокампа, полученные от крыс через одни сутки после окончания киндлинговой стимуляции. Во всех срезах производилась перерезка коллатералей Шаффера, для предотвращения влияния активности поля СА3 на эффект эпизодов повышения внеклеточной концентрации калия в поле СА1. Внеклеточная регистрация показала, что в str.pyramidale поля СА1 срезов, полученных от таких животных, уже исходно в ответ на одиночный стимул возникает множественный популяционный спайк (Рис. 3.6.2а2). Число разрядов в таком множественном спайке было достоверно выше в срезах от киндлинговых крыс, чем в срезах полученных от животных, не подвергавшихся киндлигу (2,7 ± 0,3 и 1,9 ± 0,6 у животных после киндлинга и контрольных, соответственно; p=0,008; данные получены при силе тестирующего стимула 620 μА). Этот факт указывает на то, что киндлинг in vivo приводит к генерации пачечных разрядов в пирамидных нейронах поля СА1 гиппокампа в ответ на электрическую стимуляцию, подобно тому, как это происходит при кратковременных эпизодах повышения внеклеточной концентрации калия. Если эти феномены имеют сходные механизмы, то использование эпизодов повышения калия не должно приводить к дальнейшему увеличению пачечных разрядов. Три периодических эпизода повышения K+ не приводили к дальнейшему достоверному увеличению числа разрядов в вызванном множественном спайке, регистрируемом в str.pyramidale поля СА1 срезов гиппокампа киндлинговых крыс (Рис. 3.6.2а3). Этот результат свидетельствует в пользу общности механизмов развития
Рис. 3.6.2 Окклюзия пачечных разрядов, вызываемых повышениями К+, в моделях эпилептогенеза in vitro (подпись к рисунку на следующей странице) а, Развитие пачечной активности в поле СА1 срезов гиппокампа, полученных от животных после электрического киндлинга in vivo. а1, Хроническая электрическая стимуляция гиппокампа in vivo приводила к прогрессивному развитию поведенческих судорог и электрических послеразрядов. После окончания стимуляции достигнутый уровень судорожной активности и длительности послеразрядов сохранялся как минимум неделю (1д и 1н – данные через 1 день и через 1 неделю после окончания стимуляции, соответственно). а2, Оригинальные записи, показывающие, что в пирамидных нейронах поля СА1 срезов гиппокампа животных после электрического киндлинга развивается пачечная активность. Причем последующие эпизоды повышения калия не приводили к ее усилению, что указывает, на окклюзию пачечной активности при эпизодах повышения калия пачечной активностью при эпилептогенезе. а3, Усредненные и нормированные к среднему числу спайков до воздействия данные полученные в нескольких экспериментах. На графике показана динамика изменения числа популяционных спайков в ответ на одиночную стимуляцию после 3-х повышений K+o у животных без киндлинга (-□-) и после киндлинга (-■-). Точечная линия – среднее значение числа спайков при использованной стимуляции до повышений калия, полученное в экспериментах без киндлинга. Пунктирная линия среднее значение числа спайков в экспериментах после киндлинга in vivo. Данные были нормированы к среднему до эпизодов повышения калия в каждом эксперименте. За нуль времени принята последняя регистрация перед воздействием, далее отсчет времени ведется от окончания последнего эпизода повышения K+o. б, Развитие пачечной активности в поле СА1 срезов гиппокампа, полученных от крыс линии Крушинского-Молодкиной после аудиогенного киндлинга in vivo. б1, Хроническое периодическое звуковое воздействие на животных in vivo приводило к прогрессивному развитию поведенческих судорог. После его окончания достигнутый уровень судорожной активности сохранялся как минимум неделю (1н – данные через 1 неделю после окончания звуковых воздействий). б2, Оригинальные записи, показывающие, что в пирамидных нейронах поля СА1 срезов гиппокампа крыс линии Крушинского-Молодкиной после аудиогенного киндлинга развивается пачечная активность, которой не наблюдается у животных, не подвергавшихся звуковой стимуляции. Причем последующие эпизоды повышения калия вызывают развитие пачечных разрядов у животных контрольной группы, но не приводят к ее усилению пачечных разрядов у животных после аудиогенного киндлинга. Этот результат указывает, на наличие окклюзии пачечной активности при эпизодах повышения калия пачечной активностью при аудиогенной судорожной активности. б3, Усредненные и нормированные к среднему числу спайков до воздействия данные, полученные в нескольких экспериментах. На графике показана динамика изменения числа популяционных спайков в ответ на одиночную стимуляцию после 3-х повышений K+o у животных без аудиогенного киндлинга (-○-) и после киндлинга (-●-). Точечная линия – среднее значение числа спайков при использованной стимуляции до повышений калия полученное в экспериментах без аудиогенного киндлинга. Пунктирная линия среднее значение числа спайков в экспериментах после хронических звуковых стимулов. Данные были нормированы к среднему до эпизодов повышения калия в каждом эксперименте. За нуль времени принята последняя регистрация перед воздействием, далее отсчет времени ведется от окончания последнего эпизода повышения K+o.
пачечной активности в СА1 при использовании эпизодов повышения внеклеточной концентрации калия и при киндлинге in vivo. Электрический киндлинг in vivo представляет собой экспериментальную модель эпилептогенеза, в которой судорожная активность возникает у исходно нормальных животных в результате электрической стимуляции. Это ставит под сомнение сходство его механизмов с механизмами эпилепсии, в которой значительную роль играют генетические изменения. Аудиогенная судорожная активность Одна из наиболее распространенных генетических моделей судорожной активности – аудиогенная. Аудиогенные судороги у генетически предрасположенных к ним животных связаны с патологией в слуховых путях ствола мозга (Chakravarty and Faingold 1997; Ribak et al. 1994). Однако, хроническая звуковая стимуляция может приводить к распространению судорожных разрядов из ствола мозга в лимбические структуры (Веретенников, 1996; N'Gouemo and Faingold 1997). В этом случае, чувствительность нейронов поля СА1 к экспериментальным воздействиям может измениться. Для изучения будут ли эти изменения носить характер сходный с пачечной активностью, которую вызывают повышения внеклеточной концентрации калия, мы использовали крыс линии Крушинского-Молодкиной, склонных к аудиогенным судорогам. Животные подвергались хронической звуковой стимуляции (см. Материалы и методы). При этом наблюдались прогрессирующие изменения поведенческих реакций в ответ на повторяющуюся звуковую стимуляцию: сокращение латентного периода начала судорог, изменение структуры судорог и увеличение их тяжести (Рис.3.6.2б1). Развившись, это состояние поддерживалось в течение длительного времени (не менее одной недели в проверочном эксперименте). Этот паттерн развития аудиогенных судорог получил в литературе название - аудиогенный киндлинг (Feng et al. 2001; Garcia-Cairasco et al. 1996; Kiesmann et al. 1988). В качестве контроля использовались крысы этой же линии, не подвергавшиеся хронической звуковой стимуляции. Мы регистрировали вызванные популяционные спайки в str.pyramidale поля СА1 срезов гиппокампа полученных от крыс линии Крушинского-Молодкиной: контрольных и киндлинговых. При этом у животных после киндлинга, в отличие от контрольных, наблюдались ярко выраженные множественные спайки в ответ на электрическую стимуляцию коллатералей Шаффера (число популяционных разрядов в пачке: 3,8 ± 0,7 и 1,8 ± 0,4 у животных после аудиогенного киндлинга и контрольных, соответственно, P=0,014; Рис.3.6.2б2). Затем, мы использовали три кратковременных эпизода повышения внеклеточной концентрации калия (Рис.3.6.2б3). У контрольных животных наблюдалось увеличение числа разрядов во множественном спайке, тогда как у животных после киндлинга статистически значимых изменений не наблюдалось. Этот результат свидетельствует в пользу сходства механизмов развития пачечной активности в поле СА1 гиппокампа при аудиогенной судорожной активности in vivo и при использовании эпизодов повышения внеклеточной концентрации калия in vitro.
|
||||
|
|
Последнее изменение этой страницы: 2019-04-27; просмотров: 127; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.139.69.138 (0.007 с.) |