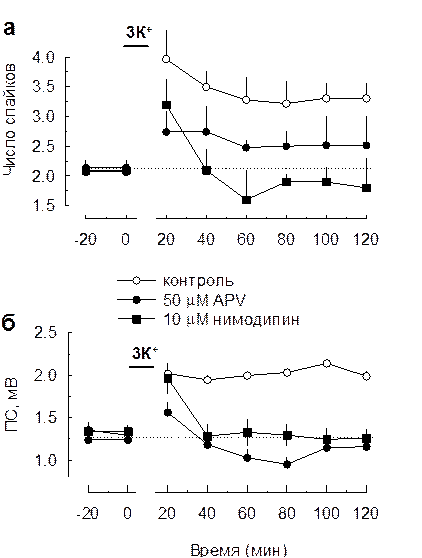Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Роль NMDA рецепторов и l-типа кальциевых каналов в повышение возбудимости пирамидных клеток и генерации пачечных разрядов
Мы исследовали какой эффект произведут три периодических эпизода повышения калия в присутствии D,L-APV (50 μM, селективного антагониста NMDA рецепторов). Добавление АPV в момент эпизодов повышения калия приводило к значительно меньшему увеличению числа популяционных спайков в ответ на электрическую стимуляцию в сравнении с контрольными экспериментами (эпизоды повышения калия без APV) (Рис. 3.6.6а). Кроме того, в присутствии APV не происходило потенциации передачи в звене пВПСП-популяционный спайк. Это результаты указывают на то, что для инициации полноценной пачечной активности в поле СА1 гиппокампа требуется повышение возбудимости клеток, в котором принимают участие NMDA рецепторы глутамата. Тем не менее, добавление APV не полностью подавило развитие множественных спайков. Эпизоды повышения калия приводили к небольшому, но статистически значимому увеличению числа популяционных разрядов в ответ на одиночный электрический стимул. Вероятным объяснением этому является то, что развитие пачечного разряда определяется не только возбуждающими механизмами, но и тормозными, а также механизмами независящими от синаптических событий. Так калиевые эпизоды приводят к деполяризации, которая влияет не только на возбуждающую синаптическую передачу, но также тормозную синаптическую передачу и внесинаптическую мембрану нейронов. До последнего времени считалось, что ГАМКергической передаче не свойственна пластичность. Однако, недавно было показано наличие посттетанических пластических изменений в ГАМКергических токах в гиппокампе (Jensen and Mody 2001; Walker et al. 2001). Причем, в зависимости от того были блокированы потенциал зависимые кальциевые каналы L-типа, тетанизация приводила либо к потенциации, либо к депрессии этих ответов (Jensen and Mody 2001). С другой стороны, L-тип кальциевых каналов может принимать участие в возбудимости внесинаптической мембраны пирамидных клеток. Поэтому, мы решили проверить, будут ли развиваться множественные разряды в поле СА1 гиппокампа, если блокировать этот тип кальциевых каналов во время эпизодов повышения калия. Добавление нимодипина (10 μМ, относительно специфического блокатора L-типа кальциевых каналов) при использовании трех периодических эпизодов повышения калия полностью подавляло как развитие пачечных разрядов, так и увеличение возбудимости пирамидных нейронов (Рис 3.6.6б). Таким образом, повышение возбудимости пирамидных нейронов определяется активацией двух различных кальциевых проводимостей. В одном случае это активация NMDA рецепторов, играющих роль в пластичности глутаматергической передачи, в другом потенциал-зависимых кальциевых каналов L-типа, принимающих участие в пластичности ГАМКергической синаптической передачи. Один их возможных механизмов заключается в том, что активация NMDA рецепторов и L-типа кальциевых каналов приводит к повышению внутриклеточного Ca2+ в пирамидных клетках. Являясь вторичным посредником, этот ион может запускать процессы фосфорилирования белков регулирующих возбудимость. Кроме этого, блокада L-типа кальциевых каналов может влиять на ГАМКергическое торможение, что в последствии отражается на активности пирамидных нейронов. Возможно, по этой причине (регуляции ГАМКергического торможения) L-тип кальциевых каналов играет более значимую роль в генерации пачечной активности, которая может являться комбинацией увеличения возбудимости пирамидных клеток и снижения их торможения. К сожалению, по причине многообразия эффектов повышения внутриклеточной
Рис. 3.6.6 Роль NMDA рецепторов и L-типа кальциевых каналов в развитии пачечной активности и увеличении возбудимости пирамидных нейронов в поле СА1 гиппокампа а, Аппликация 50 µМ APV значительно снижала увеличение числа спайков, регистрируемых в str.pyramidale поля СА1 срезов гиппокампа в ответ на одиночный стимул в str.radiatum, возникающее в результате 3-х периодических повышений внеклеточной концентрации калия (20 мМ, интервал 10 мин). Аппликация 10 µМ нимодипина полностью подавляла это увеличение. б, Аппликация как 50 µМ APV, так 10 µМ нимодипина полностью подавляла увеличение амплитуды популяционных спайков, сопровождающее увеличение числа спайков, регистрируемых в str.pyramidale поля СА1 срезов гиппокампа в ответ на одиночный стимул в str.radiatum, при использовании 3-х периодических повышений внеклеточной концентрации калия (20 мМ, интервал 10 мин).
Разбросы ± с.о.с
концентрации кальция в процессах генерации пачечной активности (Kohling et al. 2000), описать точную последовательность событий довольно трудно. Заключение Таким образом, в данном разделе работы было показано: 1. Периодические кратковременные эпизоды повышения внеклеточной концентрации калия (20 µМ; 3 эпизода по 30 сек. с интервалом 10 мин.) приводят к долговременной генерации множественных спайков в str.pyramidale поля СА1 в ответ на одиночный электрический стимул. Развитие этой активности в СА1 не зависит от событий происходящих в чувствительном к эпилептогезу поле СА3. 2. Полученные в СА1 множественные спайки не удовлетворяют всем критериям эпилептиформной активности, а частности, не регистрируются в виде спонтанных разрядов. Тем не менее, их наличие может быть частью событий в мозге, облегчающих распространение эпилептиформной активности. Это подтверждается наличием сходных по природе пачечных разрядов в поле СА1 срезов гиппокампа полученных от животных после электрического и аудиогенного киндлингов in vivo. 3. Развитие множественных спайков в поле СА1 гиппокампа сопровождается увеличением возбудимости пирамидных нейронов. Оба феномена зависят от кальциевой проводимости. Блокада L-типа потенциал-зависимых кальциевых каналов полностью подавлялет как развитие множественных спайков, так и увеличение возбудимости пирамидных нейронов. Однако, роль NMDA рецепторов является критической только для возбудимости нейронов, тогда как пачечная активность зависит лишь частично от этого типа рецепторов.
Результаты, полученные в данном разделе работы, являются важным свидетельством в пользу роли различных путей поступления кальция в клетку для генерации пачечных разрядов нейронами поля СА1 гиппокампа. Развитие пачечной активности в этом поле гиппокампа может быть важным механизмом в генерации ритмической активности: как нормальной (например, тетта ритма), так и патологической (например, эпилептиформной).
Заключение В основе функционирования ЦНС лежат три основные системы – возбуждающая, тормозная и модуляторная. Основным нейропередатчиком возбуждающей системы в головном мозге является глутамат, а тормозной - ГАМК. Модуляторные эффекты могут быть опосредованы различными системами (глутамат-, ГАМК-, моноамин‑, ацетилхолин-, пуринергической и др.) (Bouron 2001; Cherubini and Conti 2001; Khakh and Henderson 2000; Vizi 2000). Взаимодействие между этими системами обеспечивает основу для передачи, обработки и сохранения информации в мозге, а также генерации его ритмов, которые являются своего рода тактовой частотой специфичной для его областей (Boguszewicz et al. 1996; Vinogradova 2001). Нейропередатчики активируют соответствующие им рецепторы двумя способами – направленным или синаптическим, и диффузным (спилловер, volume transmission) (Agnati et al. 1995; Jansson et al. 2000; Kullmann 2000; Kullmann and Asztely 1998). Синаптическая передача передает информацию по узкому каналу цепочки синаптических переключений, тогда как диффузная изменяет активность сразу целых групп клеток за счет их внесинаптических рецепторов. Синаптическая эффективность глутаматергической передачи может меняться. Этот феномен носит название пластичности. Описаны как кратковременные, так и долговременные виды пластичности (например, LTP) (Bliss and Lomo 1973; Holscher 1999; Huang 1998; Malenka 1994; Stanton 1996). Механизмы синаптической пластичности разнообразны и включают активацию биохимических процессов в клетке (в частности, опосредованных входом кальция) (Fukunaga et al. 1996; Lisman and McIntyre 2001; Routtenberg 1985; Tokuda and Hatase 1998), перераспределение и встраивание новых постсинаптических рецепторов (Isaac et al. 1999; Man et al. 2000), изменение экспрессии определенных генов (Dragunow 1996; Walton et al. 1999) и т.д. При этом считается, что ГАМКергические синапсы не столь пластичны, по сравнению с глутаматергическими, хотя в них также обнаружены кратковременные пластические изменения (Jensen and Mody 2001). Однако, разнообразие возможных форм ионотропных ГАМКергических рецепторов указывает на более тонкую регуляцию тормозных влияний. На настоящий момент только субъединиц ГАМКергических рецепторов описано 18, а число возможных комбинаций в пентантамерной структуре несоизмеримо больше (Costa 1998; Mehta and Ticku 1999). В представленной работе нам удалось показать, что синаптические ионотропные ГАМКергические рецепторы в гиппокампе гетерогенны. Интересно отметить, что в интернейронах str.radiatum поля СА1 в генерации ТПСТ принимает участие, по всей видимости, несколько различных типов рецепторов. При этом фармакологические свойства обнаруженного нами в этих клетках типа ионотропных ГАМКергических рецепторов не являются характерными ни для одного из ранее описанных. В чем же функциональное значение различных рецепторов в одном и том же синапсе? Ответить на этот вопрос однозначно довольно сложно. ГАМКергическая передача претерпевает значительные изменения в эмбриональный и постэмбриональный периоды (Hevers and Luddens 2002; Shaw et al. 1991). Возможно, что часть ГАМКергических рецепторов присутствующих на этой стадии сохраняется и во взрослом мозге. С другой стороны, состав ионотропных ГАМКергических рецепторов может меняться при различных патологических состояниях, в частности, при эпилепсии (Nusser et al. 1998a; Schwarzer et al. 1997). Таким образом, можно предположить, что изменение пропорции функционально различных рецепторов в тормозных синапсах представляет собой форму пластичности (или защитного механизма).
|
|||||||
|
|
Последнее изменение этой страницы: 2019-04-27; просмотров: 113; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.128.190.102 (0.008 с.) |