Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Роль переднего мозга В поведении рыб.Содержание книги
Поиск на нашем сайте После изучения образного и условнорефлекторного поведения производили удаление переднего мозга у рыб. Операция проводилась способом, описанным Карамяном (13) и Холодовым (2). Передний мозг вертикальным разрезом отделяли от остальных мозговых структур и вынимали из черепной коробки. Костный дефект восстанавливали путем заливания жидкой смесью воска с вазелином (2: 1). На оперированных рыбах наблюдения велись в продолжение 2,5—3 месяцев после операции, после чего проводился морфологический контроль (Сихарулидзе, 22). Наиболее резкие изменения во внешнем поведении рыб наблюдались в продолжение 3—4 час. после операции. Сначала они неподвижно лежали на поверхности воды, после начинали плавать, покачивались из стороны в сторону, часто переворачивались на бок. Спустя 4—5 час. после операции их движения уже не отличались от нормальных.Пищу рыбы схватывали активно. К концу того же дня рыб переносили в экспериментальный аквариум. В аквариуме они ориентировались как обычно. Многочисленные опыты убедили нас, что условнорефлекторное поведение рыб не пострадало. Выработанные условные рефлексы на зрительные и звуковые раздражители вполне сохранились. Некоторое время (примерно 15—18 дней) после операции оставалась нарушенной дифференцировка условных световых сигналов (красный и желтый свет). Позднее дифференцировка восстановилась, но она была непрочной и неустойчивой, хотя надо отметить, что и у интактных рыб дифференцировка условных сигналов характеризуется неустойчивостью. После операции сильно нарушались отсроченные реакции на условные сигналы: если до операции отсроченные реакции на условные световые и звуковые раздражители были таковы, что нормальные рыбы в продолжение 8—10 сек. после сигнала правильно направлялись к сигнализируемому месту корма, то после операции при отставлении условного раздражителя (как светового, так и звукового) даже на 4—5 сек. рыбы или не выходили из обычного местонахождения пищи, или же путали направление к нему. Из этих наблюдений, между прочим, следует, что у нормальных рыб отсроченная реакция после условного разражителя наступала не в результа- те возбуждающего последействия временных связей, а благодаря образу местонахождения пищи. После удаления переднего мозга сильно пострадало образное пищевое поведение рыб, проявляемое после поедания пищи в новом месте. В нормальном состоянии рыбы направлялись прямо к этому месту и сразу после выпускания из обычного местопребывания и через 8—10 сек. После же операции они не подплывали к воспринятому месту пищевого объекта даже спустя 2—3 сек. после восприятия его. И в тех опытах, когда их подводили к пище сачком, давали поесть, а затем отводили за перегородку и сейчас же выпускали (через 4—5 сек.), они не направлялись к пищевому объекту. Если интактные рыбы избегали места раздражения после однократного электрического раздражения в новом месте, то оперированные рыбы свободно заплывали в то место, где они за несколько (4 —5) секунд получили раздражение.
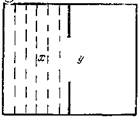
Рис. 181. Схема ящика для изучения поведения лягушки а: — отделение с проволоками на дне для электрического раздражения, у — свободное от раздражения отделение Кроме того, рыбы перестали обходить препятствия, находящиеся на пути к местонахождению пищевых объектов. У них уже не наблюдались подражательные реакции; уменьшились резко выраженные и долго сохраняющиеся реакции страха на плеск воды, открытие клетки, шум от движения экспериментатора и т. д. (Сихарулидзе, 22). Таким образом, мы убедились, что после удаления переднего мозга у рыб нарушается именно образное поведение, тогда как условнорефлекторное поведение (не считая отсроченные реакции на условные сигналы) оставалось ненарушенным. Немногочисленными исследованиями разных авторов, посвященными роли переднего мозга в условнорефлекторной деятельности рыб, было показано, что замыкание временных связей происходит в промежуточном, среднем мозге и мозжечке (1, 2) или только в мозжечке (3, 13). Итак, у костистых рыб образная память, как и память на эмоцию страха, осуществляется деятельностью переднего мозга. Условнорефлекторная же деятельность, должно быть, является функцией других отделов головного мозга.
|
|||
|
|
Последнее изменение этой страницы: 2017-02-21; просмотров: 285; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.214 (0.008 с.) |