Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Происхождение условного рефлекса на комплекс раздражении.Содержание книги
Поиск на нашем сайте
На основании приведенных наблюдений мы предположили, что в стадии дифференциации условной реакции на сложный раздражитель, когда комплекс действует положительно, а компоненты - нет, или наоборот, когда компоненты действуют положительно, а комплекс - нет, мы имеем дело с образованием новых временных связей с такими нейронами, которые приходят в возбуждение исключительно при действии данного комплекса. При этом мы исходили из того, что каждый компонент-раздражитель, как уже сказано, вызывает возбуждение в определенных нейронах коры большого мозга; в то же время в результате иррадиации возбуждения из этих возбужденных нейронов происходит повышение возбудимости в некоторых других нейронах. Если эти нейроны будут испытывать одновременно или последовательно со многих сторон такое субминимальное воздействие, то они будут приходить в возбуждение и тем самым смогут вступить во взаимодействие с теми нейронными элементами, которые возбуждаются условным и безусловным раздражениями. Понятно, если данный комплекс раздражении сочетается с безусловным раздражением, а компоненты с ним не сочетаются, тогда означенные дополнительно возбужденные нейроны в коре большого мозга должны стать начальным пунктом самостоятельных нервных связей, через которые сложный раздражитель будет производить свой собственный эффект, совершенно отличный от компонентных реакций. В вышеприведенных опытах с последовательным оборонительным рефлексом на комплексное раздражение мы также должны были иметь возникновение дополнительного активного очага под влиянием двух разных компонентов. И этот очаг должен был образовать новые временные связи на комплексное действие, так как это действие подкреплялось электрическим раздра- жением. Итак, вследствие возбуждения под влиянием комплексного раздражения таких нейронных комплексов, на которые компоненты в отдельности действуют субминималъно, сложный раздражитель производит такое физиологическое действие, которое всегда обширнее суммы физиологического эффекта компонентов. Это дополнительное возбуждение новых нейронных комплексов придает корковой деятельности объединенный целостный характер при вся-
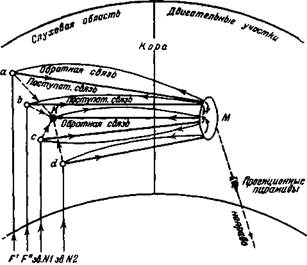
Рис. 150. Схематическая картина временных связей в отношении положительного рефлекса на комплекс из четырех звуков.
Комплекс состоял из тонов F' и F", тонвариатора Горнбостеля, звонка № 1 и звонка № 2 Компоненты в отдельности не давали рефтекса, будучи совершенно отдифференцированны, но комплекс вызывал движение.
Рис. 151. Схематическая картина временных связей, когда комплекс из тех же звуков, как на рис. 150, действовал отрицательно, не вызывая своего положительного рефлекса. Компоненты же в отдельности производили положительный рефлекс ком комплексном раздражении. Когда комплексный раздражитель сочетается с определенным жизненно важным рефлекторным действием, а компоненты не сочетаются с ним, тогда происходит образование особых временных связей с означенными дополнительно возбужденными нервными элементами. И понятно, смотря по характеру подкрепления комплексного раздражения, эти дополнительные элементы могут образовать условные связи с положительным или отрицательным внешним действием. На рис. 150 и рис. 151 дается схематическое изображение нервных вязей условного пищеводвигательного рефлекса на комплекс четырех звуков. Дополнительные нейронные комплексы (Л"), возбуждаемые комплексом звуковых раздражений, соединяются временными связями с двигательным анализатором (М), а этот последний с проекционными пирамидами через уже существующие постоянные связи.
Рис 152. Схема временных связей при осуществлении сложного пищевого передвижения На звук А собака шла к правой кормушке на звук "В — к левой На комплекс же звуков А -1- В собака оставалась на месте Один комплекс воспринимающих корковых клеток (а) соединялся временными связями с проекционными пирамидами, которые вызывали движение к правой кормушке Другой комплекс (б) был связан с другими пирамидами, воспроизводящими движение к левой кормушке Комплекс звуков А + В возбуждал двигательный нервный комплекс (К), связывавшийся с теми же проекционными пирамидами, торможение которых осуществтялось через эти связи На рис. 152 приводится схема временных связей условного пищевого рефлекса на особого рода комплекс звуков. На музыкальный звук А собака бежала к одной кормушке, а на звук В — к другой путем активации своих
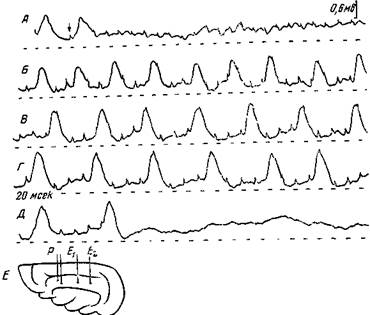
Рис. 153. Своеобразное периодическое возбуждение коры кошки при ее тетаническом раздражении Поверхностный нембуталовый сон Раздражающая пара (Р) и отводящий электрод (Iit) расположены на поверхности g suprasylvius, расстояние между ними 6 чм Интенсивность раздражения 30 в. А — вначале эффект одиночного раздражения, затем, в момент, указанный стрелкой, прикладывается раздражение с частотой 40 в сек, при этом возникают небольшие электрические потенциалы в ритме раздражения. Б — через 8 сек., В — через 9 сек.; Г — через 10 сек. от начала ргзчр^кзлия, Д — конец раздражения, Е — схема расположения электродов. Во время раздражения возникают большие потенциалы с ритмом 15—10 в сек. (Ройтбак) собственных временных связей а ж в. При одновременном действии обоих звуков собака оставалась на месте, благодаря активации временных связей с дополнительным нейронным комплексом К. Каким образом данные временные связи осуществляют положительный или отрицательный эффект, будет сказано ниже. В последнее время при электрофизиойогическом исследовании коры получены факты, которые подтверждают вышеприведенное положение о возникновении дополнительных очагов возбуждения в коре на некотором расстоянии от раздражаемого участка. Именно, Ройтбак (23) показал, что при тетаническом раздражении коры или подкоркового белого вещества сначала возникает электрическая активность по ритму раздражения на более илименее большой области, вокруг раздраженного участка (рис. 153). Но впоследствии недалеко от раздражаемого места, в ограниченном участке коры, начинают появляться иного рода электрические эффекты, по другому ритму, очевидно благодаря возбуждению новых нервных кругов из других вставочных и ассоциационных нейронов (рис. 153, Б, В и Г). Характерно, что если к такому раздражению присоединяется раздражение другого участка редкими ударами, то в означенных вторично возбужденных нервных кругах продуцируются своеобразные электрические потенциалы по ритму раздражения другого участка (рис. 154). Следовательно, данный комплекс нейронов является очагом повышенной возбудимости, легко отзывающимся на влияние побочных раздражений. Когда этот очаг находится в двигательном анализаторе, он начинает производить двигательную реакцию по ритму электрического эффекта как «спонтанно», т. е. под влиянием разных межцентральных импульсов, так и в ответ на побочное раздражение. Итак, тетаническое раздражение коры, а также многократные афферентные импульсы создают в ограниченном комплексе ассоциационных и, вставочных нейронов коры своим подпороговым действием очаги повышенной возбудимости и даже могут привести их в активное состояние. Этот дополнительно возбуждаемый комплекс нейронов с повышенной возбудимостью и должен явиться основным фактором для объединения корковой деятельности. Все дополнительные нейронные комплексы, которые служат для объединения корковой деятельности, должно быть, точно локализованы в коре большого мозга. Однако эту локализацию ни в коем случае нельзя считать прирожденно детерминированной, зависящей от наследственной организации коры большого мозга. Участие дополнительных нейронных комплексов обусловливается иррадиацией возбуждения из очага воспринимающих участков. А эта иррадиация в каждый данный момент определяется функциональным состоянием соответствующих нервных элементов, которое зависит от многочисленных условий. Следовательно, дополнительный объединяющий центр в каждом конкретном случае образуется в том или другом участке коры в зависимости от функционального состояния этих участков.
Спрашивается, где же могут быть расположены эти дополнительные, объединяющие нервные образования? Мы предположили, что объединяющая деятельность не является основной функцией тех центральных частей анализатора, которые воспринимают от рецепторов афферентную импульсацию по
Рис. 154. Периодическое возбуждение очага повышенной возбудимости в коре кошки в ответ на побочные раздражения Поверхностный нембуталовый наркоз. Раздражающие электроды (Р1 и Р2) и отводящий (8) установлены на g suprasylvms. A — пороговое раздражение (4е) через электроды Р1 с ритмом 8 в сек и присоединение сильного (30 в) раздражения через электроды Р2 с ритмом 100 в сек. Б — продолжение комбинации раздражений Р1 + Р2, при этом наступают медленные потенциалы небольшою ритма. В — прекращение рчздрчлошп Р2 при непрекращающемся раздражении Р1 Теперь на каждые два удара данного р>здртжения наблюдаются группы медленных потенциалов. Г — непосредственное продолжение записи В; Л — продолжение через 10 сек по°тс записи Г, медленные потенциалы исчезли. Е — через 40 сек., прекращение раздражения Р1. Ж — эффект сильного (25 в) раздражения Р1. 3 — схема расположения электродов (Ройтбак) специфическим путям, как, например, 17 поле в зрительном анализаторе. Объединение воспринимающих элементов должно происходить, как уже говорилось в периферических областях анализаторов, например, в отношении зрения, в полях 18 и 19, имеющих иное строение, чем 17 поле, являющееся центральным зрительным полем. Поля 18 и 19 содержат мало звездчатых клеток, но зато в них очень много малых и средних пирамид, являющихся в основном ассоциационными (см. рис. 24 и 25). На рис. 65 дана схема объединения воспринимающих нейронов зрительного анализатора, а на рис. 66 — о-бъединения этих нейронов зрительного и слухового анализаторов. При их одновременном раздражении дополнительные объединяющие нейронные комплексы должны возникать в височных ассоциационных полях, находящихся между зрительным и слуховым анализаторами. Это можно представить себе подобно тому, как это дано на рис. 66 в отношении объединения зрительных и слуховых восприятий в один образ. Из этой схемы видно, каким образом может произойти объединение зрительного и слухового анализаторов в ассоциационном поле теменной и височной областей и как каждый из этих анализаторов, будучи возбужден через соответствующие специфические афференты, может в свою очередь активировать другой анализатор путем возбуждения объединяющих нейронов в этом ассоциационном поле.
Мы здесь привели факты особого рода дифференциации условных рефлексов, когда комплексное условное раздражение вызывает рефлекс, а составляющие его компоненты, которые в отдельности не сочетались с безусловным раздражением, не дают рефлекса, или, наоборот, компоненты вызывают рефлекс, так как они сочетались с безусловным рефлексом, а комплекс, который не сочетался с ним, не дает его. При этом, когда комплекс или компоненты перестают вызывать рефлекс, они обнаруживают отрицательное действие. Мы выше пришли к выводу, что при дифференциации компонентов последние применяются без сочетания с безусловным рефлексом. Это обусловливает усиленное развитие обратных связей. Это усиливает активацию временных кругов. А так как обратные связи начинаются от ассоциационных нейронов в области проекционных пирамидных клеток, то усиленное возбуждение временных кругов должно приводить, согласно гипотезе дендритного торможения, к повышенной активации базальных дендритов этих пирамид, т. е. к усиленному анэлектротоническому действию дендритных токов на синапсы, прилегающие к этим клеткам, в частности к проекционным пирамидным клеткам комплексного рефлекса. Это явление отрицательного действия дифференцированного компонента на комплексный рефлекс проявляется и на условном оборонительном рефлексе. Как видно из рис. 147, из комплекса [сначала слабый (Сц), затем сильный (С2)] звук Ct в отдельности не дает рефлекса, но на рис. 149 он повторялся после комплекса через 5 сек. и продолжался около 15 сек., при этом рефлекса не было. Значит, звук Сг устранил последовательный комплексный рефлекс. Он наступал только после окончания компонента. Безусловно, в этом особом случае это отрицательное действие компонента должно было зависеть от усиленного развития обратных связей от двигательного анализатора к очагам компонентных действий. Вследствие этого должна была произойти усиленная импульсация из компонентных временных связей, направленная к дендритным синапсам ассоциационных и проекционных пирамидных нейронов, участвующих в производстве комплексного рефлекса. Если же компоненты производят положительные эффекты, а комплекс их отрицательный, тогда, наоборот, с дополнительно возбужденными нервными элементами комплексного раздражения обратные связи развиваются сильнее, чем с компонентными воспринимающими элементами. Так как эти обратные временные связи начинаются из области проекционных пирамид, возбуждаемых безусловным раздражением, то при возбуждении комплексных временных кругов происходит усиленная активация дендритов проекционных пирамид, которая приводит к анэлектротонической блокаде соответствующих клеточных синапсов, в частности на проекционных пирамидных клетках компонентных рефлексов. При действии компонента в отдельности дополнительный нервный центр не возбуждается и поэтому не бывает отрицательного эффекта.
Для взаимодействия комплекса и компонентов характерно, что если предъявление комплекса с отрицательным действием прекратить, но продолжать один из его компонентов, рефлекса не будет, хотя в отдельности он всегда вызывал рефлекс. Мы это объясняем тем, что в связи с возбуждением дополнительного объединяющего центра и его временных связей в них повышается возбудимость. Поэтому по прекращении комплексного раздражения они продолжают возбуждаться под влиянием компонентного раздражения, и тем самым поддерживается то отрицательное действие, которое установилось до прекращения комплексного раздражения. Аналогичный результат получится, если сначала применить компонентное раздражение с отрицательным действием, а затем заменить его комплексом с положительным действием. Можно продолжать предъявление комплекса много секунд и положительного рефлекса не будет. Надо считать, что под влиянием компонентного раздражения возбудимость повышается в соответствующих временных связях до такой степени, что по прекращении его эти временные связи продолжают активироваться усиленно под влиянием комплексного раздражения и тем самым отрицается комплексный рефлекс. Это отрицательное последействие может тянуться десятки секунд. Однако если положительный раздражитель прекратить на несколько секунд и тем обусловить понижение возбудимости во временных связях отрицательного рефлекса, то после возобновления положительного раздражения рефлекс наступит. Это взаимодействие типично для каждого случая комбинации отрицательного и положительного рефлексов. Например, на рис. 144 мы видели, что если положительный сигнал пробовать через несколько секунд после отрицательного, он вызывает рефлекс. Если же он сначала совпадает с отрицательным сигналом, а затем продолжается один десятки секунд, он рефлекса не вызывает. Надо и его прекратить на короткое время, т. е. предварительно понизить возбудимость во временных связях отрицательного рефлекса, чтобы последующий положительный сигнал произвел свое действие. Итак, при сочетании комплекса индифферентных раздражений с безусловным рефлексом, первоначально временные связи образуются со всеми компонентами, и комплексное действие равняется сумме компонентных. Но вскоре под влиянием субминимального возбуждающего действия компонентов в коре создается общий для них дополнительный очаг возбуждения. Теперь и этот дополнительный очаг образует временные связи со всеми пирамидными нейронами, возбуждаемыми безусловным раздражением. Эти связи будут по преимуществу поступательными и вызывать условную реакцию, если комплекс сочетать с безусловным раздражением, а компоненты нет. Но в это время усиленно развиваются на компонентах обратные связи. Это усиливает деятельность соответствующих нервных кругов, т. е. круговорот возбуждения в них. Сообразно исходящая из них по коллатералям импулъсация, действуя на дендриты ближайших пирамидных нейронов и вообще двигательного анализатора, может затормозить условный рефлекс на комплекс. Если же компоненты сочетаются с безусловным раздражением, а комплекс нет, тогда на дополнительном очаге усиленно развиваются обратные связи. Сообразно исходящая из нервных кругов этого очага импулъсация будет активировать дендриты пирамидных нейронов в двигательном анализаторе и вызывать торможение компонентных рефлексов.
|
|||||||||||
|
|
Последнее изменение этой страницы: 2017-02-21; просмотров: 256; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.168.68 (0.013 с.) |