Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Характеристика условного рефлекса на комплекс раздражений.
Выше рассматривалась корковая деятельность при оборонительных или пищевых условных рефлексах на один простой раздражитель, на один какой-либо звук или на свет электрической лампы, т. е. путем сочетаний такого простого раздражителя с безусловным рефлексом. С новыми явлениями корковой деятельности мы сталкиваемся, когда вырабатываем условный рефлекс на сложный раздражитель, на комплекс разных звуков или на сложную фигуру.В этом случае мы имеем, с одной стороны, связывание частей сложного раздражителя с безусловным рефлексом, а с другой, объединение частей сложного раздражителя в одно целое и связывание этого целого с безусловным рефлексом. В лаборатории Павлова эта проблема была поставлена с самого же начала. Впервые Зелёный в 1907 г. (36) уделил этой проблеме большое внимание. Он образовал условный слюнный рефлекс на комплекс звуков и затем изучал взаимоотношение между данными компонентами и комплексом. Зелёный установил, что первоначально и компоненты дают рефлекс, подобно комплексу, но только слабее комплекса. Если компоненты одинаковы по интенсивности, то они вызывают одинаковые рефлексы, которые почти вдвое слабее, чем целый комплекс; если же один компонент сильнее другого, то в отдельности он дает больший эффект, чем слабый. Разница наблюдается как в скрытом периоде, так и в величине секреции. Далее, Зелёный установил, что с течением времени компоненты перестают давать рефлексы, в то время как комплекс, воздействующий сейчас же после них, вызывает его без ослабления. Этой стадии полной дифференциации компонентов предшествует такой период, когда испытание компонентов отражается на величине комплексного рефлекса: последний ослабевает. Зелёный подробно изучил отношение компо- нентных рефлексов друг к другу и установил, в общем, такие же отношения, как между однородными условными рефлексами (36). Но эти фактические результаты не получили дальнейшего развития, не привели к выяснению физиологических основ объединяющей деятельности коры большого мозга. Все последующие в этом направлении работы школы Павлова, а также Бехтерева касаются, как и работа Зеленого, констатирования объединяющей деятельности, или, как теперь принято называть в этих лабораториях, синтетической способности, наряду с аналитической (Бабкин, 37; Палладин, 38; Фурсиков, 39; Иванов-Смоленский, 40; Яковлев, 41; Лунина, 42).
Рис. 145. Третий день образования рефлекса на левой передней ноге на чесание спины и звук тонвариатора 290 гц Собака Белый 35-е сочетание Во время чесания (Чес) лишь слабое подергивание левой передней (средняя кривая) и правой передней ноги (нижняя кривая) и начало ориентировочной реакции (верхняя кривая). Во время звука (290 гц) сгибается левая передняя нога, потом наступает общее движение с более сильным поднятием левой ноги После звука, спустя несколько секунд, электрическое раздражение левой передней ноги (верхний сигнал с обозн. 8) Иванов-Смоленский (40) приходит к выводу об образовании из комплекса разных раздражителей одного «целого, слитного раздражения» и отсюда заключает о том, что «в коре больших полушарий происходит процесс интегрирования (сложного синтеза)» (1928). Но он ограничивается только одним констатированием факта. Условнорефлекторная деятельность коры большого мозга на комплекс раздражителей изучалась Беритовым совместно с Брегадзе по методу свободных движений (43). Было произведено нами обширное исследование, в котором подтвердились данные из школы Павлова; кроме того, было открыто много других фактов. Мы первоначально изучали оборонительный рефлекс собаки на комплекс двух раздражений — сначала чесание спины, а затем давался звук тонвариатора Штерна. Безусловное электрическое раздражение производилось спустя несколько секунд после звука. В другой серии опытов применяли один и тот же звук органной трубы, но в двух интенсивностях: сначала слабый звук, а затем сильный. Хотя тон был один, надо рассматривать слабое и сильное звучание как различные по качеству раздражения. Это, прежде всего, потому, что оба тона слабого и сильного звука не производили одинаковое физиологическое действие. Прислушиваясь к данным звукам, мы улавливали заметную разницу в тембре звука. В обеих сериях каждый компонент длился 5—10 сек., а затем после перерыва 2—3 сек. давался другой компонент такой же длительности. Лишь после этого через 5—10 сек. прикладывалось электрическое раздражение к ноге, которое вызывало обычную оборонительную реакцию: общее передвижение, визг, учащенное дыхание и поднятие раздражаемой ноги.
После ряда подкреплений комплекс начинал вызывать оборонительные движения, но первоначально они наступали еще во время действия компонентов, а компоненты в отдельности также вызывали оборонительный рефлекс. Особенно активен был всегда второй компонент. Так как основное, безусловное раздражение производилось после условного, то рефлекс вскоре стал последовательным как в ответ на комплекс, так и на компоненты. Например, при образовании комплексного условного оборонительного рефлекса на чесание
Рис. 14G. Образование последовательного рефлекса на левой передней ноге на чесание и на звук 290ец Та гке собака, что и на рис 145 Третий день работы В опыте 1 — производится только кратковремен ный звук (290 гц), который вызывает двойной последовательный реф текс, сначала сгибается левая пе рсдняя нога (средняя кривая), за тем правая (нижняя кривая) Ори снтировочная реакция наступает сейчас же по прекращении звука (верхняя кривая) В опыте 2— сначата производится чесание (Чес), затем звук Теперь также рефлекс явчяется двойным и следует сейчас же после прекращения звука Но нч чтот раз реакция на каждой ноге протекает интенсивнее, чем в предыдущем опыте при отсутствии че сания Ориентировочная реакция также значительно си шнес, чем в опыте 1 спины 10—15 сек., а затем звук 290 гц — 15—20 сек. условный рефлекс уже после 35 сочетаний стал последовательным: нога поднялась спустя несколько секунд после звука (рис. 145). Но что характерно, если испытать один звук — 290 гц, то вызывается последовательный рефлекс, но более слабый, чем после комплекса (рис. 146). При образовании оборонительного рефлекса на комплекс звуков — сначала слабый, а затем тот же звук органной трубы большей интенсивности — электрическое раздражение лапы производилось спустя несколько секунд после звуков. После первых сочетаний рефлекс наступал еще во время звуков, главным образом во время второго, более сильного звука. Затем рефлекс стал последовательным. Каждый звук в отдельности вызывал рефлекс, но второй, более сильный, вызывал большее сгибание лапы (рис. 147). После систематического сочетания комплекса с электрическим раздражением рефлекс стал наступать в конце комплексов или после них, т. е. стал последовательным. Проба компонентов в отдельности также давала последовательный рефлекс, он был сильнее на второй компонент, чем на первый. Но в общем оба компонентных рефлекса были слабее комплексного. В дальнейшем оба компонента в отдельности не вызывали рефлекса. Более того, они не вызывали и в том случае, если один и тот же компонент повторялся два раза или сначала действовал сильный компонент, а потом слабый (рис. 148). Были испробованы также звуки органной трубы на полтора тона ниже обычного. Они также не давали рефлекса или давали значительно слабее (Беритов и Топурия, 44). В общем при анализе фактов мы пришли к заключению, что в первый период образования оборонительного рефлекса на комплекс компоненты вызывали его через свои самостоятельные поступательные временные связи
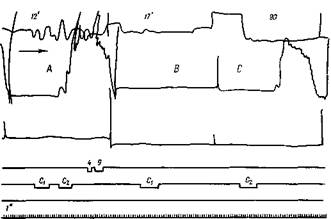
Рис. 147. Образование последовательного рефлекса на комплекс звуков
Собака Джек. Слабый звук — С,; сильный — С2 (органной трубы) Четвертый день работы. Спокойное состояние животного В опыте А дается обычный комплекс. Сильный условный рефлекс наступает еще во время второго звука. В опыте В испытывается звук Ci, который не дает эффекта, а в опыте С испытывается компонент С,- Он производит значительный последовательный эффект. Верхняя кривая— от головы, средняя — от левой передней ноги, нижняя — от правой задней ноги
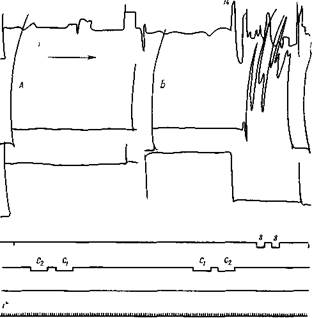
Рис. 148. Дифференциация рефлекса Собака Джек. В опыте А — дается необычный комплекс С2 — Ci, который не дает рефлекса. Есть лишь ничтожное ориентировочное движение головы (верхняя кривая). В опыте Б— спустя 6 мин. после опыта А, дается обычный комплекс, который производит последовательный рефлекс. В обоих опытах до раздражения собака висит на лямках с вытянутыми вперед ногами, но не спит
Рис. 149. Влияние звука d на последовательный рефлекс на комплекс С, — С2 Собака Джек. В опыте А обычное сочетание. Поднятие ноги началось спустя 2 сек после звука. В опыте Б — сейчас же после обычного комплекса применяется компонент d, рефлекс начался по окончании Сг, приблизительно через 10 сек. с двигательным анализатором, но они были слабее комплексного рефлекса. Последний равнялся сумме компонентных. Но впоследствии, после многократных испытаний без подкрепления электрическим раздражением они перестали вызывать рефлекс, тогда как комплекс продолжал вызывать его. Произошла дифференциация, благодаря сильному развитию обратных связей, как это указывалось выше в отношении рецепторной дифференциации условных рефлексов. Это обусловило отрицательное действие компонентов на положительный комплексный рефлекс. Если один из компонентов присоединялся после комплекса, последовательный рефлекс, соответствующий комплексу, не наступал. Он появлялся через несколько секунд после присоединенного компонента (рис. 149). Совершенно очевидно, что в стадии дифференциации комплексного рефлекса нельзя рассматривать комплексное действие суммой компонентных. Безусловно здесь мы имеем дело со сложным физиологическим действием, о котором подробно будет сказано ниже при рассмотрении другой серии опытов, выполненной на собаках более обширно по методике свободных движений, на основе пищевого поведения. 1. При одновременном действии комплекса из четырех звуков (тон 340 ец и тон 680 гц, звонок № 1 и звонок другого тембра № 2) животное отводилось от лежанки к кормушке и там подкармливалось. В этот момент звуки прекращались и после закрытия кормушки животное отводилось обратно. После нескольких подкармливаний не только комплекс звуков, но и каждый компонент вызывал пищевое поведение — собака шла к кормушке. Но так как при изолированном применении отдельных компонетов собака не подкармливалась, она вскоре перестала при их применении ходить к кормушке. Собака бежала к кормушке только при комплексе звуков. Более того, если компоненты комплекса применялись не одновременно, а включались последовательно, один за другим, собака не шла к кормушке. Она не шла к кормушке даже в том случае, если сначала применяли один компонент, а затем сразу присоединяли остальные три (Беритов и Брегадзе, 43). Итак, в то время как комплекс звуков оказывает на животное положительное влияние, вызывает передвижение к кормушке, дифференцированные компоненты действуют на него отрицательно, устраняя это движение к кормушке.
2. В другой серии опытов водили собаку к кормушке только применяя отдельные компоненты звукового комплекса. Когда собака сама стала бе- гать к кормушке при действии компонентов, стали пробовать комплекс «одновременно все компоненты» без подкармливания. Собака вначале при этом бегала к кормушке, но пищи не получала. Довольно скоро она перестала реагировать на комплекс звуков, а продолжала бегать к кормушке только при изолированном применении отдельных компонентов (Беритов и Брегадзе, 43). 3. Вырабатывалось пищевое поведение на один музыкальный тон к кормушке 1, которая находилась налево от собаки, а на другой тон — к кормушке 2, расположенной направо от нее. При одновременной пробе этих звуков собака сначала бегала то к одной, то к другой кормушке. Но так как в этих случаях собака не подкармливалась, то вскоре добились того, что при одновременном действии этих звуков она оставалась на лежанке, в то время как при изолированном действии того или другого компонента собака шла к соответствующей кормушке. Более того, мы обнаружили, что когда комбинация звуков не вызывает пищевого поведения, она действует отрицательно на это поведение. А именно, если вскоре после пробы комплекса применяли один компонент, он не вызывал движения к кормушке (Беритов иБрегадзе, 43). 4. Аналогичные результаты были получены при образовании индивидуального пищевого поведения на сложную световую фигуру. И в этом случае отдельные компоненты ее переставали вызывать пищевое поведение. То же наблюдалось и при образовании пищевого поведения на комплекс световых и звуковых раздражений. При подкармливании собаки только на этот комплекс— свет и звук, применяемые в отдельности компоненты не вызывали передвижения к кормушке (Брегадзе, 44). 5. Было выработано также пищевое поведение на два или три последовательных звуковых раздражений, например, на do3, sol3 и do4. Каждый звук продолжался 5 сек., а по окончании через несколько секунд собаку отводили к кормушке. Вначале пищевое поведение наступало во время действия звуков. Однако животное подкармливалось только по окончании их. Поэтому после нескольких или многократных испытаний собака шла к кормушке только после прекращения действия комплекса. Звуки, применяемые в отдельности, перестали вызывать пищевое поведение. Более того, испытание аналогичного ряда звуков из верхних или нижних октав не вызывало пищевого поведения с самого начала или после нескольких проб без сочетания с едой (Беритов, 2; Брегадзе, 72).
Мы здесь получили точно такого же рода факты, как при образовании последовательного условного оборонительного рефлекса на комплекс слабый — сильный звук одного тона органной трубы. На основании приведенного фактического материала мы пришли к выводу, что при образовании пищевого поведения на комплекс одновременных или последовательных раздражений оно вначале вызывается не комплексом раздражений, как данным целостным воздействием, а всеми входящими компонентами самостоятельно. При этом реакции на компоненты были тем значительнее и прочнее, чем сильнее их физиологическое действие. Следовательно, в начале образования положительного пищевого поведения на сложный раздражитель каждый компонент вызывает передвижение животного к кормушке через свои собственные временные связи, а весь комплекс — через временные связи всех входящих в него компонентов. Впоследствии, когда компоненты перестают уже вызывать пищевое поведение, мы не можем рассматривать положительный рефлекс как сумму компонентных реакций, поскольку эти последние проявляют отрицательное действие на пищевое поведение, вызываемое комплексом раздражений. Это особенно хорошо видно в том случае, когда пищевое поведение вырабатывается на отдельные компоненты, а весь комплекс применяется без подкармливания. Выработанная при этом отрицательная реакция на комплекс не может быть признана за сумму положительных компонентных эффектов.
|
|||||||||||
|
|
Последнее изменение этой страницы: 2017-02-21; просмотров: 231; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.216.251.37 (0.023 с.) |