Схема патогенеза механической желтухи
Содержание книги
- Полиоловый путь превращения глюкозы
- Анаэробный распад глюкозы (гликолиз). Этапы. Гликолитическая оксидоредукция. Регуляция. Физиологическое значение анаэробного распада глюкозы. Баланс энергии.
- Аэробное окисление глюкозы. Изложить анаэробную фазу.
- Изменение скорости реакций цтк и причины накопления кетоновых тел при некоторых состояниях
- Мобилизация гликогена (гликогенолиз)
- Схематичное расположение дефектных ферментов при различных гликогенозах
- Включение глицерина в синтез глюкозы
- Высшие жирные кислоты, структура, свойства, биологическая роль. Понятие о полиненасыщенных жирных кислотах.
- Гликолипиды - рецепторные молекулы
- Липиды пищи, их характеристика. Суточная потребность в липидах.
- Полный ферментативный гидролиз триацилглицерола
- Нарушения переваривания и всасывания пищевых жиров.
- Ресинтез триацилглицеринов в стенке кишечника.
- Формирование кетоновых тел. Причины и последствия кетоза.
- Удлинение цепи жирных кислот
- Реакции синтеза таг из фосфатидной кислоты
- Холестерин, строение, свойства, основные этапы синтеза и его регуляция.
- Пищевые продукты - источники белков. Нормы белка в питании. Биологическая ценность белков. Понятие об азотистом балансе.
- Переваривание белков в желудке, характеристика ферментов. Роль соляной кислоты.
- Изменение кислотности в желудке
- Переваривание белков и полипептидов в кишечнике. Характеристика протеиназ.
- Возможные пути превращений аминокислот
- Схема реакции трансаминирования
- Декарбоксилирование аминокислот. Гистамин, серотонин и другие биогенные амины.
- Появление аммиака в клетках непрерывно
- Реакции глюкозо-аланинового цикла (выделен рамкой).
- Взаимосвязь обмена серина, глицина, метионина и цистеина
- Синтез креатина и креатинина, креатинфосфат - дополнительный источник энергии мышечного сокращения.
- Патология белкового обмена. Белковое голодание. Причины и последствия.
- Врожденные нарушения обмена некоторых аминокислот (фенилкетонурия, алкаптонурия, цистиноз и цистинурия).
- Распад нуклеиновых кислот в тканях. Катаболизм пуриновых оснований.
- Реакции катаболизма пуриновых нуклеотидов
- Источники атомов пуринового кольца
- Биосинтез пиримидиновых нуклеотидов в тканях.
- Нарушения порфиринового обмена. Порфирии.
- Этапы метаболизма билирубина в организме
- Схема патогенеза механической желтухи
- Патологические формы гемоглобина
- Белки сыворотки крови. Отдельные представители. Биологические функции.
- Гамма-глобулины - защитные белки
- Единицы измерения активности ферментов
- Роль почек в регуляции водно-солевого обмена. Вазопрессин, альдостерон, ренин-ангиотензиновая система.
- Физико-химические свойства мочи в норме и ее химический состав.
- У мочи есть и другие особенности
- Патологические компоненты мочи. Клиническое значение анализа мочи.
- Эндогенная вода. Компартментализация жидкостей в организме. Значение воды в организме.
- Биологическая роль, регуляция уровня.
- Биологическая роль, регуляция уровня. Ионы кальция необходимы для передачи нервного импульса
- Регуляция синтеза и секреции. Активируют: Гипокальциемия повышает гидроксилирование витамина D по С1 в почках через
- Железо, его концентрация в сыворотке крови. Биологическая роль. Определение железа в сыворотке крови.
Биохимическими маркерами синдрома холестаза (в "чистом" виде, без повреждения гепатоцитов) являются:
1. Гипербилирубинемия за счёт связанного (прямого) билирубина.
2. В моче высокий уровень билирубина (коричневый цвет, цвет темного пива) и снижено количество стеркобилина, уробилина нет.
3. В кале практически отсутствует стеркобилин (обесцвеченность, серовато-белое окрашивание).
4. Повышение в сыворотке крови активности ферментов, специфичных для желчных канальцев – щелочная фосфатаза (желчный изофермент), 5′-нуклеотидаз а, γ-глутамилтранспептидаза.
5. Уменьшение концентрации альбуминов и увеличение содержания α2-, β- и γ-глобулинов в сыворотке крови (протеинограмма для механической желтухи).
Паренхиматозная желтуха
Паренхиматозная (печеночно-клеточная) желтуха – причиной может быть нарушение на всех трех стадиях превращения билирубина в печени:
· извлечение билирубина из крови печеночными клетками,
· конъюгирование билирубина,
· АТФ-зависимая секреция в желчь.
Наблюдается при различных формах гепатитов (вирусные, токсические) и иных поражениях печени.
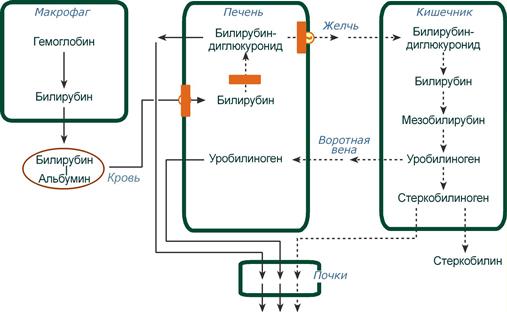
Схема патогенеза паренхиматозной желтухи
У младенцев вариантами паренхиматозной желтухи являются физиологические желтухи новорожденных и недоношенных:
· физиологическая желтуха,
· желтуха, вызываемая молоком матери и т.п.
Наследственные желтухи печеночного происхождения представляют собой синдромы Жильбера-Мейленграхта, Дубина-Джонсона, Криглера-Найяра.
Биохимическими маркерами синдрома цитолиза являются:
1. Гипербилирубинемия за счёт обеих фракций билирубина – прямой и непрямой. Их концентрация в крови возрастает из-за одновременного нарушения секреции в желчь и увеличения проницаемости мембран клеток печени.
2. В моче имеется билирубин (коричневый цвет, цвет черного чая), появляется уробилин.
3. В кале стеркобилин снижен или в норме.
4. Повышение в сыворотке крови активности ферментов, специфичных для гепатоцитов – ЛДГ-5, АЛТ, АСТ, γ-глутамилтранспептидаза, глутаматдегидрогеназа.
5. Уменьшение концентрации альбуминов и увеличение содержания β- и γ-глобулинов в сыворотке крови (протеинограмма для гепатитов).
Биосинтез гема.
Гем является небелковой частью многих гемопротеинов:
· гемоглобин (до 85% общего количества гема организма), локализованный в эритроцитах и клетках костного мозга,
· миоглобин скелетных мышц и миокарда (до 17%),
· цитохромы дыхательной цепи,
· ферменты цитохромоксидаза, цитохром P 450, гомогентизатоксидаза, миелопероксидаза, каталаза и глутатионпероксидаза, тиреопероксидаза и т.д. – менее 1%.
Строение и синтез гема
Гем – структура, включающая в себя порфириновое кольцо (состоящее из 4 пиррольных колец) и иона Fe2+. Железо связывается с порфириновым кольцом двумя координационными и двумя ковалентными связями.
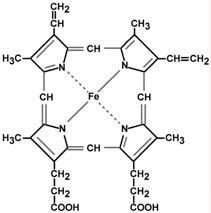
Строение гема
Синтез гема в основном идет в предшественниках эритроцитов, в клетках печени, почек, слизистой кишечника и в остальных тканях. Первая реакция синтеза с участием δ-аминолевулинат-синтазы (греч. δ - "дельта") происходит в митохондриях. Следующая реакция при участии аминолевулинатдегидратазы (порфобилиноген-синтазы) протекает в цитозоле, здесь из двух молекул δ‑аминолевулиновой кислоты образуется циклический порфобилиноген (монопиррол).
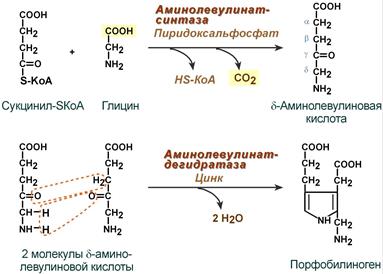
Синтез порфобилиногена
После синтеза порфобилиногена четыре его молекулы конденсируются в гидроксиметилбилан, который далее превращается в уропорфириноген типа I и уропорфириноген типа III. В синтезе обоих видов порфиринов принимает участие уропорфириноген I-синтаза, в образовании уропорфириногена III дополнительно принимает участие фермент уропорфириноген III-косинтаза.
Судьба обоих типов уропорфириногена двояка: они могут окисляться до уропорфирина (на рисунке не показано) или декарбоксилироваться до копропорфириногена соответствующего типа.
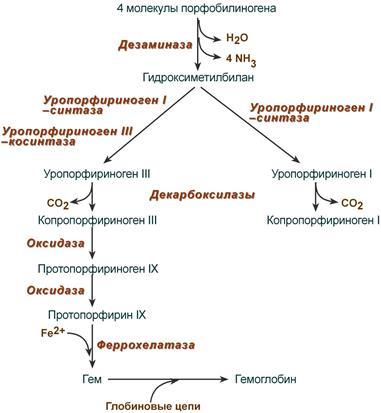
|






