Полиоловый путь превращения глюкозы
Содержание книги
- Витамин Д. Биологическая роль.
- Обобщенная схема эффектов кальцитриола
- Витамин Е. Биологическая роль.
- Строение двух форм витамина К
- Три механизма передачи сигнала в зависимости от вида мембранных рецепторов
- Гормоны щитовидной железы. Роль тироксина и тиреокальцитонина в обмене веществ.
- Схема реакций синтеза тиреоидных гормонов
- Паратгормон. Роль в обмене веществ.
- Регуляция синтеза и секреции. · ангиотензин II, выделяемый при активации ренин-ангиотензиновой системы,
- Регуляция синтеза и секреции. Активируют: АКТГ, обеспечивающий нарастание концентрации кортизола в утренние часы
- Регуляция синтеза и секреции. Активируют: кортиколиберин при стрессе (тревога, страх
- Гормоны задней доли гипофиза вазопрессин, структура и его роль в осмотическом концентрировании мочи.
- Мужские половые гормоны. Синтез, механизм клеточного действия. Влияние на обмен веществ.
- Строение женских половых гормонов
- Регуляция синтеза и секреции. Секреция инсулина происходит постоянно, и около 50% инсулина
- Два механизма действия инсулина
- Реакции, связанные с активацией MAP-киназного пути
- Нарушение обмена веществ при инсулиновой недостаточности.
- Взаимосвязь различных видов обмена. Регуляция метаболизма.
- Общие понятия о биологическом окислении. Значение работ советских ученых в открытии и развитии учения о биологическом окислении. АТФ - универсальная форма энергии в клетке.
- Комплекс. НАДН-КоQ-оксидоредуктаза
- Понятие о метаболических путях. Общие и специфические пути катаболизма углеводов, жиров, аминокислот.
- Роль оксалоацетата в метаболизме
- Строение некоторых производных моносахаридов
- Гетерополисахариды. Отдельные представители. Биологическая роль.
- Полиоловый путь превращения глюкозы
- Анаэробный распад глюкозы (гликолиз). Этапы. Гликолитическая оксидоредукция. Регуляция. Физиологическое значение анаэробного распада глюкозы. Баланс энергии.
- Аэробное окисление глюкозы. Изложить анаэробную фазу.
- Изменение скорости реакций цтк и причины накопления кетоновых тел при некоторых состояниях
- Мобилизация гликогена (гликогенолиз)
- Схематичное расположение дефектных ферментов при различных гликогенозах
- Включение глицерина в синтез глюкозы
- Высшие жирные кислоты, структура, свойства, биологическая роль. Понятие о полиненасыщенных жирных кислотах.
- Гликолипиды - рецепторные молекулы
- Липиды пищи, их характеристика. Суточная потребность в липидах.
- Полный ферментативный гидролиз триацилглицерола
- Нарушения переваривания и всасывания пищевых жиров.
- Ресинтез триацилглицеринов в стенке кишечника.
- Формирование кетоновых тел. Причины и последствия кетоза.
- Удлинение цепи жирных кислот
- Реакции синтеза таг из фосфатидной кислоты
- Холестерин, строение, свойства, основные этапы синтеза и его регуляция.
- Пищевые продукты - источники белков. Нормы белка в питании. Биологическая ценность белков. Понятие об азотистом балансе.
- Переваривание белков в желудке, характеристика ферментов. Роль соляной кислоты.
- Изменение кислотности в желудке
- Переваривание белков и полипептидов в кишечнике. Характеристика протеиназ.
- Возможные пути превращений аминокислот
- Схема реакции трансаминирования
- Декарбоксилирование аминокислот. Гистамин, серотонин и другие биогенные амины.
- Появление аммиака в клетках непрерывно
Отдельное значение путь имеет в семенных пузырьках, которые синтезируют фруктозу для семенной жидкости. Сперматозоиды, находящиеся здесь, используют фруктозу в качестве основного энергетического субстрата, но при переходе в женские половые пути переключаются на глюкозу. Считается, что использование фруктозы повышает стабильность и предотвращает преждевременное разрушение акросомы спермия.
Клиническая значимость полиолового пути проявляется в инсулиннезависимых тканях, в которые глюкоза поступает неконтролируемо по градиенту концентрации – эндотелий, хрусталик глаза, клетки клубочков почек, но в первую очередь, нейроны. Путь активируется при гипергликемиях, когда поток глюкозы в клетки увеличивается и скорость синтеза сорбитола резко возрастает.
Накопление сорбитола в нейронах подавляет синтез циклического спирта инозитола, входящего в состав фосфолипидов и используемого при передаче гормонального сигнала, и снижает активность Na +,K + -АТФазы, что нарушает проведение нервного импульса. Избыток в клетке осмотически активных сорбитола и фруктозы удерживает воду, изменяет форму клеток и их функциональную активность.
Избыточная работа альдозоредуктазы приводит к расходованию НАДФН в клетках, что подавляет ряд процессов, например
· восстановление глутатиона, используемого в антиоксидантной системе клеток и в метаболических процессах,
· реакцию синтеза вазодилататора оксида азота (NO),
· нейтрализация аммиака в реакции восстановительного аминирования α-кетоглутарата,
· микросомальное окисление.
Судьба глюкозы
После активации (фосфорилирования) глюкоза, в зависимости от условий и вида клетки, превращается по различным направлениям:
· часть глюкозы обязательно используется в энергетическом обмене, она сгорает в реакциях аэробного и анаэробного гликолиза для синтеза АТФ,
· при достаточно большом количестве глюкоза запасается в виде гликогена, к синтезу гликогена способны большинство тканей,
· в гепатоцитах (при высокой концентрации), в миоцитах и в адипоцитах глюкоза перенаправляется на синтез триацилглицеролов,
· в печени и иных тканях глюкоза используется для синтеза холестерола,
· при определенных условиях часть глюкозы идет в реакции пентозофосфатного пути, в котором образуются рибозо-5-фосфат и НАДФН,
· некоторая доля глюкозы используется для синтеза глюкозаминов и далее структурных или иных гетерополисахаридов.
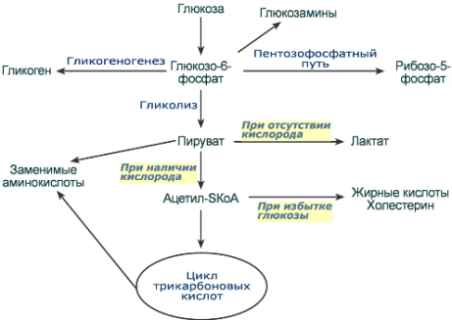
Реакции превращения глюкозы в клетке
Гексокиназа - первый фермент метаболизма глюкозы
Наличие глюкозы в клетке обеспечивается, в первую очередь, облегченной диффузией ее из крови в цитозоль при участии специальных транспортных белков - глюкозных транспортеров (ГлюТ).
Активация глюкозы
После перемещения через мембраны глюкоза в цитозоле немедленно фосфорилируется ферментом гексокиназой, в связи с чем фермент образно называют " ловушка глюкозы ". Фосфорилирование глюкозы решает несколько задач:
· фосфатный эфир глюкозы не в состоянии выйти из клетки, так как молекула отрицательно заряжена и отталкивается от фосфолипидной поверхности мембраны,
· наличие заряженной группы обеспечивает правильную ориентацию молекулы в активном центре фермента,
· уменьшается концентрация свободной (нефосфорилированной) глюкозы в клетке, что способствует диффузии новых ее молекул из крови.
Дефосфорилирование глюкозы осуществляется глюкозо-6-фосфатазой. Этот фермент есть только в печени и почках. В эпителии канальцев почек работа фермента связана с реабсорбцией глюкозы. В гепатоцитах фермент необходим, когда печень поддерживает гомеостаз глюкозы в крови.
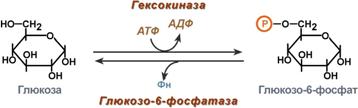
|






