Аэробное окисление глюкозы. Изложить анаэробную фазу.
Содержание книги
- Витамин Е. Биологическая роль.
- Строение двух форм витамина К
- Три механизма передачи сигнала в зависимости от вида мембранных рецепторов
- Гормоны щитовидной железы. Роль тироксина и тиреокальцитонина в обмене веществ.
- Схема реакций синтеза тиреоидных гормонов
- Паратгормон. Роль в обмене веществ.
- Регуляция синтеза и секреции. · ангиотензин II, выделяемый при активации ренин-ангиотензиновой системы,
- Регуляция синтеза и секреции. Активируют: АКТГ, обеспечивающий нарастание концентрации кортизола в утренние часы
- Регуляция синтеза и секреции. Активируют: кортиколиберин при стрессе (тревога, страх
- Гормоны задней доли гипофиза вазопрессин, структура и его роль в осмотическом концентрировании мочи.
- Мужские половые гормоны. Синтез, механизм клеточного действия. Влияние на обмен веществ.
- Строение женских половых гормонов
- Регуляция синтеза и секреции. Секреция инсулина происходит постоянно, и около 50% инсулина
- Два механизма действия инсулина
- Реакции, связанные с активацией MAP-киназного пути
- Нарушение обмена веществ при инсулиновой недостаточности.
- Взаимосвязь различных видов обмена. Регуляция метаболизма.
- Общие понятия о биологическом окислении. Значение работ советских ученых в открытии и развитии учения о биологическом окислении. АТФ - универсальная форма энергии в клетке.
- Комплекс. НАДН-КоQ-оксидоредуктаза
- Понятие о метаболических путях. Общие и специфические пути катаболизма углеводов, жиров, аминокислот.
- Роль оксалоацетата в метаболизме
- Строение некоторых производных моносахаридов
- Гетерополисахариды. Отдельные представители. Биологическая роль.
- Полиоловый путь превращения глюкозы
- Анаэробный распад глюкозы (гликолиз). Этапы. Гликолитическая оксидоредукция. Регуляция. Физиологическое значение анаэробного распада глюкозы. Баланс энергии.
- Аэробное окисление глюкозы. Изложить анаэробную фазу.
- Изменение скорости реакций цтк и причины накопления кетоновых тел при некоторых состояниях
- Мобилизация гликогена (гликогенолиз)
- Схематичное расположение дефектных ферментов при различных гликогенозах
- Включение глицерина в синтез глюкозы
- Высшие жирные кислоты, структура, свойства, биологическая роль. Понятие о полиненасыщенных жирных кислотах.
- Гликолипиды - рецепторные молекулы
- Липиды пищи, их характеристика. Суточная потребность в липидах.
- Полный ферментативный гидролиз триацилглицерола
- Нарушения переваривания и всасывания пищевых жиров.
- Ресинтез триацилглицеринов в стенке кишечника.
- Формирование кетоновых тел. Причины и последствия кетоза.
- Удлинение цепи жирных кислот
- Реакции синтеза таг из фосфатидной кислоты
- Холестерин, строение, свойства, основные этапы синтеза и его регуляция.
- Пищевые продукты - источники белков. Нормы белка в питании. Биологическая ценность белков. Понятие об азотистом балансе.
- Переваривание белков в желудке, характеристика ферментов. Роль соляной кислоты.
- Изменение кислотности в желудке
- Переваривание белков и полипептидов в кишечнике. Характеристика протеиназ.
- Возможные пути превращений аминокислот
- Схема реакции трансаминирования
- Декарбоксилирование аминокислот. Гистамин, серотонин и другие биогенные амины.
- Появление аммиака в клетках непрерывно
- Реакции глюкозо-аланинового цикла (выделен рамкой).
- Взаимосвязь обмена серина, глицина, метионина и цистеина
В аэробном процессе пировиноградная кислота превращается в ацетил-SКоА (реакции ПВК-дегидрогеназы) и далее сгорает в реакциях цикла трикарбоновых кислот до СО2 (реакции ЦТК).
Общее уравнение аэробного окисления глюкозы:
C6H12O6 + 6 O2 + 32 АДФ + 32 Фнеорг → 6 CO2 + 44 H2О + 32 АТФ
Аэробным называется окисление биологических субстратов с выделением энергии, протекающее при использовании кислорода в качестве конечного акцептора водорода в дыхательной цепи. В качестве доноров водорода выступают восстановленные формы коферментов (НАДН, ФАДН2 и НАДФН), образующиеся в промежуточных реакциях окисления субстратов.
Аэробное дихотомическое окисление глюкозы является основным путём катаболизма глюкозы в организме человека и может происходить во всех органах и тканях. В результате этих реакций глюкоза расщепляется до СО2 и Н2О, а выделяющаяся энергия аккумулируется в АТФ. В этом процессе можно условно выделить три стадии:
1. превращение глюкозы в 2 молекулы пирувата в цитоплазме клеток (специфический путь распада глюкозы);
2. окислительное декарбоксилирование пирувата с образованием ацетил-КоА в митохондриях;
3. окисление ацетил-КоА в цикле Кребса в митохондриях.
15.3.2. На каждом этапе процесса происходит образование восстановленных форм коферментов, которые окисляются ферментными комплексами дыхательной цепи с образованием АТФ путём окислительного фосфорилирования. Коферменты, образующиеся на второй и третьей стадиях аэробного окисления глюкозы, подвергаются непосредственному окислению в митохондриях. В то же время НАДН, образующийся в цитоплазме в реакциях первой стадии аэробного окисления, не способен проникать через митохондриальную мембрану. Перенос водорода с цитоплазматического НАДН в митохондрии происходит при помощи специальных челночных циклов, основным из которых является малат-аспартатный челночный механизм. Цитоплазматический НАДН восстанавливает оксалоацетат в малат, который проникает в митохондрию, где окисляется, восстанавливая митохондриальный НАД; в цитоплазму оксалоацетат возвращается в виде аспартата (рисунок 15.3).
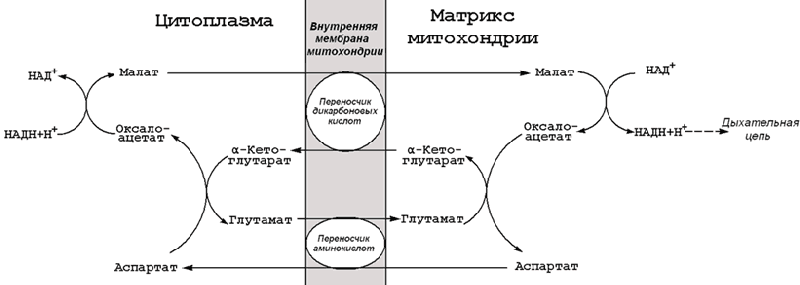
Рисунок 15.3. Малат-аспартатный челночный механизм.
Продукция АТФ в реакциях аэробного дихотомического окисления происходит также в трёх реакциях субстратного фосфорилирования – две из них в гликолизе, третья в цикле Кребса на уровне сукцинил-КоА. Полный энергетический баланс аэробного окисления глюкозы представлен на рисунке 15.4.

Рисунок 15.3. Энергетический баланс аэробного окисления глюкозы.
Аэробное окисление глюкозы. Челночные механизмы.
Челночные системы
Так как сама молекула НАДН через мембрану не проходит, то существуют специальные системы, принимающие атомы водорода от НАДН в цитоплазме и отдающие их в матриксе митохондрий. Эти системы получили название челночные системы.
Определены две основные челночные системы – глицеролфосфатная и малат-аспартатная.
На основании наличия алкогольдегидрогеназы во многих тканях, в том числе и в нервной, дискутируется вопрос о существовании этанол-ацетальдегидной челночной системы, однако однозначных экспериментальных доказательств пока не получено.
|






