Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Чувствительные нервные окончания – сенсорные рецепторыСодержание книги
Поиск на нашем сайте
Общая характеристика Рецепторы – конечные специализированные образования, предназначенные для трансформации энергии различных видов раздражителей в специфическую активность нервной системы. Изначально потенциальным раздражителем для животного является любое изменение внешней среды, так как задачей рецепции и является оценка её неоднородности. Как уже отмечалось, эту задачу в ходе эволюции взяли на себя, естественно, клетки наружного слоя гипотетического предка многоклеточных, непосредственно с этой средой контактирующие (см. разд. 1.2). Доказательством этому является формирование тканей нервной системы из эктодермы. Как уже отмечалось выше, в основе возникновения специфических рецепторных клеток лежит функциональная асимметрия покровных тканей, непосредственно контактирующих как с внешней, так и внутренней средой. У ряда клеток этих тканей соответственно гипертрофируется либо апикальный, либо базальный конец клетки. Поэтому, несмотря на всё многообразие рецепторов, исходной для всех рецепторных клеток моделью является жгутиковая клетка. Основные же молекулярные механизмы рецепции были «разработаны» ещё в процессе эволюции одноклеточных.
Рецепторные клетки отличаются от других в двух отношениях: 1) для них энергия раздражителя служит лишь стимулом к запуску процессов, совершаемых за счёт потенциальной энергии, которая накоплена вследствие обменных реакций в самой клетке; 2) рецепторная клетка обладает на выходе энергией, обязательно передаваемой другим клеткам, которые сами не способны воспринимать энергию данного внешнего воздействия. Основной структурной единицей большинства рецепторных аппаратов является клетка, снабжённая подвижными волосками или ресничками – «антеннами». Они действуют подобно усилителю по отношению к воспринимаемым раздражителям и участвуют в трансформации раздражителя в нервную сигнализацию. Организация рецепторных нервных аппаратов многоклеточных разнообразна. Это обуславливается тем, что: 1) каждый тип рецепторов приспособляется к восприятию сигнала одного типа, специфического; 2) в рецепторных нейронах и специализированных для этой цели клетках других тканей происходит преобразование энергии внешнего стимула в энергию нервного импульса или рецепторного потенциала.
Структурно рецепторы подразделяются на 1) свободные и 2) несвободные. 1). У свободных терминальные ветвления периферического отростка чувствительного нейрона лежат непосредственно в эпителии или соединительной ткани. 2). Несвободные рецепторы помимо терминальных ветвлений периферического отростка чувствительного нейрона имеют специальные (глиальные) клетки. Практически все окончания этого типа – механорецепторы; инкапсулированные механорецепторы имеют оформленную соединительнотканную капсулу.
По отношению к раздражителю рецепторные клетки делятся на две группы: 1) первичночувствующие и 2) вторичночувствующие. 1). В первичночувствующих (первичных) рецепторных клетках действие адекватного раздражителя осуществляется непосредственно периферическим отростком сенсорного нейрона, который, таким образом, непосредственно, первично встречается с раздражителем. Он находится на периферии, а не в центральной нервной системе, и представляет собой преобразованный биполярный нейрон, на одном полюсе которого – дендрит с ресничкой или дендритным отростком, на другом – центральный отросток – аксон. Такие рецепторы впервые обнаруживаются у кишечнополостных и являются основным универсальным типом рецепторных элементов, с которым связаны все виды рецепции у беспозвоночных. У позвоночных они представлены тканевыми рецепторами и проприорецепторами, а также терморецепторами и обонятельными клетками (см. ниже). 2). Во вторичночувствующих (вторичных) рецепторных клетках между окончанием сенсорного нейрона и точкой приложения раздражителя располагается дополнительная специализированная клетка ненервного происхождения. Возникающее в ней возбуждение передаётся через синапс на сенсорный нейрон – следовательно, он возбуждается не первично внешним стимулом, а опосредованно, благодаря воздействию рецептирующих клеток. Последние не имеют периферических и центральных отростков; восприятие стимула у них осуществляется с помощью жгутикообразных выростов. Это боковая линия, волосковые клетки внутреннего уха, рецепторные клетки вкусовых луковиц, фоторецепторы глаза и др.
По принципу взаимодействия с раздражителем рецепторы подразделяются на а) контактные и 2) дистантные. Как следует из названия, дистантные, в отличие от первых, позволяют животному ориентироваться в пространстве, не вступая в прямой контакт с источником раздражения (органы зрения, обоняния, слуха, гравитации, терморецепторы).
В биологическом плане по источнику импульса рецепторы многоклеточных можно подразделить на 1) экстеро- и 2) интерорецепторы, то есть осуществляющие восприятие состояния соответственно а) внешней и б) внутренней среды. У экстерорецепторов больше выражена специализация – высокая избирательность к адекватному раздражителю. Они могут реагировать и на неадекватные стимулы, но лишь на очень интенсивные. Поэтому экстерорецепторы принято считать мономодальными. Среди интероцепторов тоже есть мономодальные образования. Однако большинство - полимодальны, то есть способны реагировать на несколько разных раздражителей – например, термические, химические и механические. Интерорецепторы Итак, эти рецепторы служат для восприятия специфических раздражений из внутренней среды организма. Примером могут служить так называемое тельце Фатер-Пачини (фатерпачиниевое) (рис. 23) и рецепторы растяжения мышц млекопитающих.
Рис. 23. Схема строения тельца Фатер-Пачини (по Отели и др.): 1 – слоистая капсула, 2 – внутренняя колба, 3 – дендрит чувствительной нервной клетки, Эти тельца находятся в соединительной ткани кожи и различных органов. Они относятся к числу инкапсулированных рецепторов. Их размер – 0,5-1,0 мм. Тельце образовано терминальными разветвлениями дендрита чувствительного нейрона, заключённого в капсулу, сформированную специализированными фиброцитами, коллагеновыми волокнами и клетками глии. К тельцу подходит толстое миелинизированное нервное волокно. Генерация нервного импульса осуществляется в результате деформации концевого участка дендрита, эффект которой усиливает сложная капсула; непосредственно в генерации импульса последняя не участвует. Рецепторы растяжения в поперечнополосатых мышцах позвоночных представлены у двух типов мышечных волокон – медленных и быстрых (см. разд. 2.3.3.2.2). На каждом из них – два типа окончаний: а) дендритов чувствительных клеток и б) нервно-мышечные синапсы возбуждающих нейронов. Экстерорецепторы Как следует из названия, это рецепторы, обеспечивающие поступление информации из внешней среды - то есть зрительные, слуховые, вкусовые, обонятельные, болевые, температурные, тактильные и пр. По типу воспринимаемых факторов можно выделить прежде всего механо-, фото-, хемо- и терморецепторы. Механорецепторы. Этот тип рецепции возникает из первичной чувствительности к механическим воздействиям всей поверхности клеточной мембраны одноклеточных. У многоклеточных рецепторную функцию различных тканей и органов в основном осуществляют рецепторные клетки нересничного типа, тогда как в акустико-латеральной системе рецепторными клетками являются волосково-реснитчатые. Механорецепцию осуществляют периферические отделы соматических, скелетно-мышечных, слуховых и вестибулярных сенсорных систем, а также боковой линии.
Простейшим примером механорецепторов являются чувствительные волоски насекомых. У них кутикулярный волосок (см. разд. 1.2.2.1.3) контактирует со специализированным дендритом биполярной чувствительной клетки, тело которой расположено в гиподерме; благодаря такому взаимодействию при деформации щетинки и возникает нервный импульс. Относящиеся к этой группе рецепторы гравитации (равновесия) у подавляющего большинства животных строятся по одной схеме (рис. 24). В любом органе равновесия имеется так называемая пробная масса – массивное подвижное внеклеточное образование разного состава и происхождения, способное перемещаться при изменении положения организма в гравитационном поле и тем самым механически воздействовать на разные группы окружающих рецепторных клеток. Второй обязательный компонент органа равновесия - чувствительная антенна, представленная видоизменёнными жгутиками и микроворсинками. Они образуются нервными или особыми чувствительными клетками, генерирующими нервный импульс (нервная клетка) или рецепторный потенциал (чувствительная клетка).
Рис. 24. Схема строения органа гравитации (по www. 1.jpg): 1 – пробная масса; 2 - желеобразная масса; 3 – рецепторная клетка с выростами; 4 – обонятельный нерв; 5 – эпителий (а и б – положение относительно вектора гравитации). Органы гравитации в процессе эволюции претерпевают большие усложнения, благодаря чему высшие животные получают возможность не только ориентироваться в статическом гравитационном поле, но и анализировать угловые ускорения при перемещении тела. Фактически по аналогичному типу устроены органы слуха и сейсмосенсорной системы, где роль пробной массы выполняет либо специальная жидкость (в первом случае), либо окружающая вода (во втором). Хеморецепторы. Чувствительны к действию химических агентов. У наземных животных они образуют периферические отделы обонятельной и вкусовой сенсорных систем, тогда как для водных различия в этих понятиях не имеют явного смысла. Участвуют в оценке химического состава внутренней среды и связаны с работой висцерального анализатора. Типичными хемоэкстерорецепторами являются органы вкуса и обоняния (рис 25). У высших животных они достигают сложной структурной организации. В основе восприятия химического раздражения в дистантном хеморецепторе (органе обоняния) и контактном хеморецепторе (органе вкуса) лежит один и тот же механизм. Его основа - взаимодействие молекул различных веществ со специфическими белковыми молекулами, вмонтированными в специализированные мембраны соответствующих первично - или вторичночувствующих клеток. Строение же соответствующих чувствительных органов у различных животных довольно разнообразно.
Рис. 25. Схема строения обонятельного рецептора (по A. Frestein, 2001): Фоторецепторы. Представлены цилиарными рецепторами, то есть производными от клеток со жгутиками, и рабдомерами, у которых жгутик отсутствует, а собственно фоторецепторная часть образована совокупностью микровилл (микровыростов плазмолеммы). У всех многоклеточных они представляют собой клетки со специализированными дендритами. У позвоночных это так называемые палочки и колбочки (рис. 26).
ампулированные и бугорковые электрочувствительные рецепторные органы.
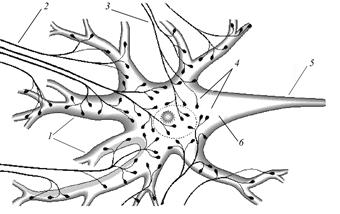
Болевые (ноцицептивные). Наряду со специализированными, болевые стимулы могут воспринимать и другие рецепторы. Синапсы Главные функциональные процессы, происходящие в нейроне – генерация, проведение и передача нервного импульса на другой нейрон или иную дифференцированную клетку. Место контакта двух клеток, где и происходит передача импульса, называется синапсом. По тому, с какой на какую части нейрона передаётся импульс, синапсы подразделяют на аксодендритические, аксо-аксональные, денродендритические, аксосоматические, («сома» - тело, в данном случае – нейрона). Количество синапсов, приходящихся на один нейрон, различно. У высших позвоночных их может быть несколько сотен (рис. 27).
Рис. 27. Синаптические окончания на двигательной нервной клетке
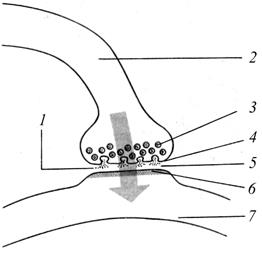
Механизм действия синапсов различен. В настоящее время известны два типа синапсов: 1) химические - передающие нервный импульс с помощью специального вещества-посредника (медиатора) строго в одном направлении; 2) электрические (электротонические) - передающие импульс в обоих направлениях без химических посредников; 3) смешанные. Химические синапсы В структуру х имического синапса входят следующие компоненты (рис. 28): а) пресинаптическая мембрана (с которой передаётся импульс), б) синаптическая щель (между пре- и постсинаптической мембранами) и в) постсинаптическая мембрана (на которую импульс передаётся).
Рис. 28. Схема типичного синапса (между нейронами А. и Б) (по Албертсу Б. Эти образования фиксированы круговой синаптической пластинкой и сетью десмосом между пре- и постсинаптической мембранами. Пресинаптический полюс – расширенное окончание аксона с митохондриями и синаптическими пузырьками. В последних содержатсявещества- медиаторы, то есть «передатчики», «посредники» возбуждения. Большинство нейромедиаторов – аминокислоты и их производные (табл. 10). Таблица 10
|
|||||||||||
|
|
Последнее изменение этой страницы: 2017-02-10; просмотров: 210; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.189.171.137 (0.012 с.) |

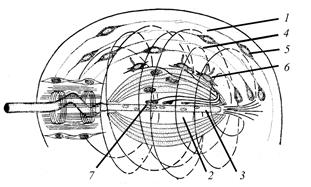
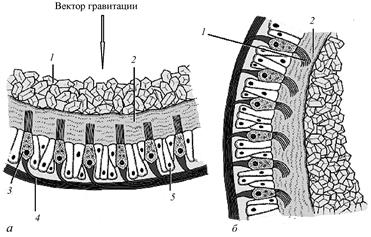
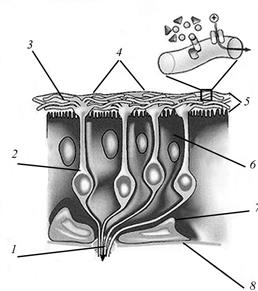
 Рис.26. Схема строения фоторецепторов позвоночных животных (по Заварзину А. А., 2000): 1 – наружный сегмент, 2 – внутренний сегмент, 3 – скоп-ление митохондрий, 4 – отростки клеток пигментного эпителия, 5 – ядро, 6 – синаптическая зона, 7 – масляная капля.
Рис.26. Схема строения фоторецепторов позвоночных животных (по Заварзину А. А., 2000): 1 – наружный сегмент, 2 – внутренний сегмент, 3 – скоп-ление митохондрий, 4 – отростки клеток пигментного эпителия, 5 – ядро, 6 – синаптическая зона, 7 – масляная капля.