Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Функциональная классификация нейронов.
В зависимости от задач исследования даже с точки зрения их функционирования нейроны могут быть классифицированы по-разному. По месту в составе рефлекторной дуги различают нейроны: 1) чувствительные – воспринимают внешний стимул и преобразуют его в нервный импульс (большинство таких клеток находится в органах чувств, в нервных узлах); 2) ассоциативные (вставочные) - в основном в составе центральной нервной системы; 3) эффекторные (моторные, двигательные) - передают нервный импульс на рабочий орган. (В качестве разновидности последних нередко рассматривают группу нейросекреторных клеток, которые при получении нервного импульса выделяют вещества, регулирующие работу многих тканей организма). По направлению возбуждения различают нейроны: а) афферентные - обеспечивают продвижение нервного импульса к телу клетки (в том числе это чувствительные нейроны разных модальностей, восходящих путей) и б) эфферентные нейроны двигательных путей и трактов - обеспечивают продвижение импульса к периферии. Также существует классификация нейронов по числу отростков (см. ниже). Строение нейронов (Приложение, рис. 22, 23) Размеры нейронов – от 4 до 140 мкм. В них различают: 1) тело нейрона (ядросодержащая часть, перикарион); 2) отростки; 3) концевые аппараты (нервные окончания) – структуры, которыми заканчиваются отростки. Тело нейрона имеет различную форму (круглую, овальную, пирамидную, грушевидную, веретеновидную). Для поддержания его структуры служит развитый цитоскелет, образованный сетью нейрофибрилл. Они, в свою очередь, сформированы нейротрубочками диаметром около 24 нм (с ними связывают внутриклеточный транспорт) и нейрофиламентами диаметром 10 нм (в отростках нейрофибриллы лежат параллельно оси отростка). Ядро (как правило, крупное, с деконденсированным мелкодисперным хроматином и крупным ядрышком) – располагается обычно в центре клетки. В нейронах есть все органоиды общего назначения. При этом очень хорошо развита гранулярная эндоплазматическая сеть с многочисленными крупными плоскими параллельными цистернами - тельцами Ниссля, формирующими так называемый тигроид. Последний имеется также в основании дендритов; зона отхождения аксона – аксонный холмик – свободна от него. Количество тигроида резко снижается при переутомлении или при перерезке аксона, а при его регенерации вновь возрастает.
В гранулярной эндоплазматической сети нейрона происходит синтез белков (необходимых для поддержания массы отростков), медиаторов (для обеспечения работы синапсов), а также белков, необходимых для обеспечения обучения и запоминания. На протяжении всей жизни клетки её синтетический аппарат работает с очень большой нагрузкой, обеспечивая функциональную деятельность нейрона и постоянную внутриклеточную регенерацию.
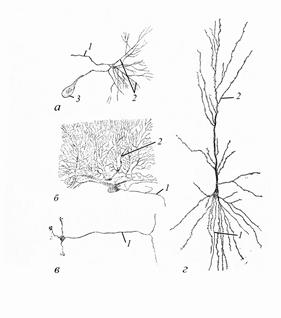
От перикариона отходят отростки. Функционально различают аксоны (отростки, которые проводят импульс от тела нейрона) и дендриты (проводят возбуждение к перикариону). Аксон - длинный отросток, как правило, не ветвящийся, но образующий концевые разветвления, в которых содержатся синаптические пузырьки. Его объём может достигать 99% суммарного объёма нейрона. Любой нейрон имеет лишь один аксон. Дендриты – ветвящиеся (за что и получили своё название) отростки, заканчивающиеся вблизи тела нейрона. Проксимальная часть дендрита – продолжение перикариона, и потому имеет аналогичные внутренние структуры. Дендриты интегрально образуют до 95% рецепторной поверхности (рецептивного поля) нейрона. В своей цитоплазме они содержат те же органоиды, что имеются и в области перикариона. Дендриты растут и подвергаются ретракции в ответ на различные воздействия, включая активность нейрона, действие различных веществ (в том числе лекарственных препаратов), повреждение соседних нейронов. Дендриты, в отличие от аксонов, способны генерировать не только потенциалы действия, возникающие по принципу «всё или ничего», но и градуальные генераторные потенциалы. Число дендритов у нейрона – от нуля до нескольких. Длина отростков нейронов может достигать 1м и более. По числу отростков нейроны классифицируют на (рис. 20): 1) униполярные – имеют один отросток (аксон); 2) биполярные – имеют два отростка (аксон и дендрит); более широко распространены у беспозвоночных животных; у позвоночных представлены преимущественно специализированными рецепторными клетками органов зрения и обоняния и некоторыми ассоциативными нейронами;
Рис. 20. Псевдоуниполярные и мультиполярные нейроны беспозвоночных и позвоночных животных (по А. А. Заварзину, 2000): а – двигательная клетка грудного ганглия личинки стрекозы, б – клетка Пуркинье, в – клетка-зерно,
3) близки по строению к биполярным псевдоуниполярные нейроны, у которых имеется один общий вырост, который затем разветвляется на аксон и дендрит (в онтогенезе закладываются два отростка, которые, сближаясь, образуют общий ствол); 4) мультиполярные – один аксон и несколько дендритов (большинство клеток центральной и периферической нервной системы). Это наиболее разнообразный тип нейронов. У позвоночных животных ими является большинство ассоциативных и эффекторных нейронов соматической и все нейроны вегетативной нервной системы. По-видимому, в отдельную группу можно выделить широко распространённые у беспозвоночных различных групп униполярные нейроны с особой дифференцировкой рецепторных областей на отростке или его разветвлениях, где можно выделить зоны: а) проксимальный (вставочный) сегмент – связующее звено между трофической (перикарион) и рабочей (отростки) частями клетки; б) рецептивный сегмент – аналогичен «обычным» дендритам; в) аксон. Впрочем, перечисленными разновидностями всё многообразие нейронов не исчерпывается. По аксону от тела клетки к окончанию движется цитоплазма, формируя в аксонном транспорте (аксотоке) три основных потока: - медленный (1-3 мм в сутки) – перемещение трофических белков, ферментов, микропузырьков; - быстрый (5-10 мм в час) – перемещение нейросекреторных гранул; - промежуточный – движение митохондрий и лизосом. Дендритный транспорт (движение цитоплазмы от тела клетки по дендритам) – в основном обеспечивает перемещение белков, необходимых для расщепления медиаторов. Его скорость близка скорости быстрого аксотока. Имеет место и ретроградный (обратный) ток цитоплазмы, благодаря которому перикарион получает информацию о состоянии нервных окончаний. Большая часть отростков нейронов покрыта специальными оболочками, образованными клетками олигодендроглии (см. ниже).
В нервной системе высших многоклеточных процессы репродукции нервных клеток обычно стойко блокированы. Однако у этих животных в постнатальном гистогенезе, а у низкоорганизованных животных в течение всей жизни иногда происходят процессы соматической полиплоидизации, за счёт которой достигается интенсификация внутриклеточного метаболизма. В отличие от самих нейронов, которые способны только к внутриклеточной физиологической регенерации, отростки нейронов способны к репаративной регенерации. При этом регенерация осуществляется за счёт проксимальной части повреждённого отростка со скоростью около 1-4 мм в сутки; в этом процессе большое значение имеют клетки нейроглии. Нейроглия Из описанного выше следует, что основные клетки нервной системы - нейроны - имеют целый ряд своеобразных биологических особенностей: необычное ядерно-плазменное отношение, высокий уровень метаболической активности, большую функциональную подвижность специфических структур и др. Поэтому для нормального функционирования они нуждаются в особой среде со специальными механическими, трофическими, защитными, изолирующими свойствами, механизмами. Эту задачу и выполняет глия, составляющая значительную часть компонентов нервной системы (например, она составляет почти половину объёма головного мозга).
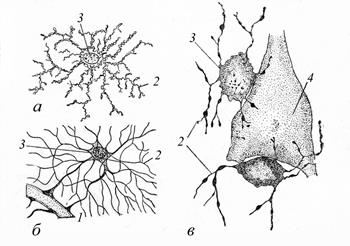
Нейроглия возникает на ранних этапах эволюции нервной системы (у кишечнополостных не обнаружена). В силу разнообразного строения органов нервной системы клетки глии весьма разнообразны по строению и функциям. Наибольшего разнообразия нейроглия достигает у позвоночных животных. Морфологически все глиальные клетки этих животных можно (с известными оговорками) разделить на две большие категории: макро- и микроглию. Микроглия. Её клетки имеют небольшие размеры, неправильную форму, многочисленные ветвящиеся отростки, множество лизосом. Они являются фагоцитами; их активация сопровождает различные заболевания. Макроглия (рис. 21), происходящая из нервного зачатка, в свою очередь, подразделяется на: 1) древнюю эпендимную (иногда её выделяют в отдельную группу), 2) астроцитарную и 3) олигодендроглию.
Рис. 21. Клеточные элементы макроглии в сером веществе центральной нервной системы млекопитающих (по А. А. Заварзину, 2000): 1). Эпендимная глия (эпендима, глиоэпителиальная ткань). Её клетки (цилиндрической формы) образуют пласт, сходный с эпителием, так как эти ткани имеют сходные функции. Эпендима выстилает центральный канал спинного и желудочки головного мозга. Её клетки имеют хорошо развитые реснички на апикальной поверхности и многочисленные пузырьки в цитоплазме. Также в составе этой ткани присутствуют и иные (нересничные) клетки. Эпендима выполняет разграничительную функцию, участвует в секреции, регуляции состава ликвора (цереброспинальной жидкости), в его перемещении. Эта ткань является ярким примером конвергентного сходства тканей разного происхождения (в данном случае – с мерцательным эпителием), обусловленного сходством выполняемой функции (в данном случае локомоции). 2). Астроциты - «звёздчатые клетки», названные так за свою отростчатую форму. Их отростки отходят от тела клетки в разных направлениях, оплетая нейроны, сосуды, клетки эпендимы и др.
В свою очередь астроциты можно подразделить на: а) волокнистые - с длинными, слабо или совсем не ветвящимися отростками (присутствуют в белом веществе мозга); б) протоплазматические - с многочисленными короткими ветвящимися отростками (находятся в сером веществе). Функции астроцитов разнообразны; основными из них являются: а) транспорт питательных веществ, газов и метаболитов от кровеносных сосудов и к ним; б) регуляция химического состава межклеточной жидкости, в том числе концентрации нейромедиаторов; в) изоляция рецептивных поверхностей нейронов; г) участие в пролиферации и замещении погибших нейронов; д) фагоцитоз и экспрессия иммунных реакций; е) синтез веществ, способствующих росту аксонов и др. 3). Олигодендроглия. Её клетки, как правило, более мелкие. В сером веществе мозга олигодендроциты находятся в непосредственном контакте с перикарионами и отростками нейронов; в белом веществе они расположены рядами между нервными волокнами. Так называемые шванновские клетки (леммоциты) входят в состав миелиновых (мякотных) и безмиелиновых (безмякотных) нервных волокон, образуя их оболочки, и рассматриваются как аналоги олигодендроцитов. Каждая такая клетка покрывает один аксон. Между собой эти клетки образуют щелевые контакты. Шванновские клетки - так называемые миелинобразующие. Миелин – компактная структура из мембран, которая формируется описанным выше путём. 70 % массы миелина составляют липиды; важными компонентами являются также белки. Именно благодаря миелину так называемое белое вещество центральной нервной системы (то есть скопление нервных волокон) имеет характерный цвет, отличающий его от серого (строение и образование нервных волокон описано ниже). Рассматриваемые клетки выполняют ряд функций: опорную, защитную, трофическую, разграничительную (по отношению к нейронам); обеспечивают ускорение проведения нервного импульса по отросткам нейронов; участвуют в регенерации нервных волокон. Нервные волокна (Приложение, рис. 24, 25) Отросток нервной клетки (называемый в данном случае осевым цилиндром), покрытый оболочкой из леммоцитов – это и есть нервное волокно. Обычно различают два типа волокон: 1) безмякотные (безмиелиновые) - находятся преимущественно в составе вегетативной нервной системы; проводят нервный импульс по цитолемме осевого цилиндра со скоростью 1-2 м/с; 2) мякотные (миелиновые) – представлены преимущественно в составе центрального и периферического отделов соматической нервной системы; благодаря миелинизации скорость проведения нервного импульса в них гораздо выше – 5-120 м/с. При участии леммоцитов нервные волокна формируются следующим образом. При образовании более простого по строению безмиелинового волокна, возникающего в онтогенезе раньше, вдоль отростка нейрона, вплотную прилегая к нему, выстраивается цепочка леммоцитов. Далее последние в месте контакта прогибаются и постепенно полностью охватывают отросток подобно муфте. При этом цитолемма шванновской клетки, сомкнувшись над отростком, образует складку (так называемый мезаксон – участок сдвоенной цитолеммы) – на которой осевой цилиндр как бы подвешен внутри леммоцита. На этом образование безмиелинового волокна заканчивается. Часто в одну клетку погружается с разных сторон несколько (5-20) отростков нейрона, формируя волокно «кабельного типа».
Формирование миелиновых волокон (рис. 22) начинается подобно безмиелиновым. При этом вращательное движение осевого цилиндра по часовой стрелке и движение леммоцита в противоположном направлении приводят к удлинению и постепенному накручиванию 10 и более слоёв мезаксона вокруг осевого цилиндра. В миелине на некотором расстоянии друг от друга имеются светлые полоски – насечки миелина («насечки Шмидта-Лантермана»). Они являются следствием движения цитоплазмы леммоцита, раздвигающей в данном месте «завитки» мезаксона. Через некоторые интервалы (от нескольких микрон до нескольких миллиметров) волокно имеет участки, лишённые миелинового слоя. Это - зоны контакта двух соседних леммоцитов (так называемые межузловые перехваты или перехваты Ранвье). Участок между двумя соседними перехватами называется межузловым сегментом.
Рис 22. Мякотное нервное волокно (по J. Cleland, 2001): а – последовательность формирования; б – сформированное волокно. 1- аксон; 2 - шванновская клетка; 3 - перехват Ранвье. Собираясь в пучки, нервные волокна формируют нервы. При этом между отдельными волокнами располагаются прослойки рыхлой соединительной ткани – эндоневрий; отдельные группы волокон объединены периневрием; наконец, нерв в целом окружён эпиневрием. Нервные окончания Общая характеристика Нервные окончания – это те структуры, в которых, несмотря на название, как з аканчиваются, так и начинаются основные процессы, обеспечивающие общую интеграцию организма и связь его с внешней средой. Нервные окончания представляют собой концевые аппараты отростков нервных клеток. Их можно подразделить на: 1) эффекторные – как следует из названия, располагающиеся на аксонах эффекторных нейронов; 2) чувствительные (рецепторные, афферентные) – окончания дендритов чувствительных нейронов; 3) синаптические, которые находятся на аксонах рецепторных нейронов, на дендритах эффекторных клеток, на любых отростках вставочных нейронов, а также входят в состав межнейрональных синапсов (см. ниже), осуществляющих связь нейронов друг с другом. Эффекторное нервное окончание – это окончание аксона эффекторного нейрона на мышце или секреторной клетке. Примером может служить нервно-мышечное окончание на скелетной мышце – так называемая «моторная бляшка». В зоне её формирования нервное волокно теряет миелиновую оболочку и разветвляется, погружаясь при этом в мышечное волокно, прогибая его плазмолемму. Это пресинаптический полюс моторной бляшки. Он содержит синаптические пузырьки с медиатором (ацетилхолином) и митохондрии. Участок мышечного волокна в зоне контакта с нервным окончанием формирует постсинаптический полюс. Здесь теряется поперечно-полосатая исчерченность мышечного волокна, содержится много митохондрий. Плазматическая мембрана, которая имеет рецепторы к медиатору, формирует многочисленные складки, дающие вторичные синаптические щели (см. ниже). В этой области накапливается фермент для разрушения медиатора - холинэстераза. Между двумя полюсами образуется синаптическая щель шириной около 50 нм. Когда нервный импульс, идущий по отростку нейрона, достигает пресинаптического полюса, медиатор выделяется в синаптическую щель, воздействует на холинорецепторы постсинаптической мембраны, и вызывает волну деполяризации плазмолеммы мышечного волокна, что и влечёт за собой сокращение последнего. После чего медиатор разрушается холинэстеразой. На гладких мышцах и секреторных клетках эффекторы устроены проще: они представлены кольцевыми утолщениями аксона с синаптическими пузырьками.
|
|||||||||
|
|
Последнее изменение этой страницы: 2017-02-10; просмотров: 763; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.9.141 (0.033 с.) |