Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Гамк - ергическая нейромедиаторная система и физиологически активные вещества, нарушающие её функционированиеСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Министерства здравоохранения Российской Федерации
Факультет военного обучения
ГАМК - ЕРГИЧЕСКАЯ НЕЙРОМЕДИАТОРНАЯ СИСТЕМА И ФИЗИОЛОГИЧЕСКИ АКТИВНЫЕ ВЕЩЕСТВА, НАРУШАЮЩИЕ ЕЁ ФУНКЦИОНИРОВАНИЕ
Учебное пособие «Рекомендуется Учебно-методическим объединением по медицинскому и фармацевтическому образованию вузов России в качестве учебного пособия для студентов медицинских вузов»
Москва
Авторы: Бабенко Олег Васильевич – начальник Факультета военного обучения Московской медицинской академии им. И.М. Сеченова, кандидат медицинских наук, доцент, полковник медицинской службы Гладких Вадим Дмитриевич – профессор кафедры «Медицинской профилактики и защиты» Факультета военного обучения Московской медицинской академии им. И.М. Сеченова, доктор медицинских наук, полковник медицинской службы Кирьянов Владимир Владимирович – начальник учебной части – заместитель начальника кафедры «Медицинской профилактики и защиты» Факультета военного обучения Московской медицинской академии им. И.М. Сеченова, подполковник медицинской службы
ГАМК – ергическая нейромедиаторная система и физиологически активные вещества, нарушающие её функционирование /Под ред. О.В. Бабенко. - М.: ММА им. И.М. Сеченова, 2004. - с. В настоящем учебном пособии изложены вопросы метаболизма гамма-аминомасляной кислоты (ГАМК) как основного медиатора торможения нервной системы. С учётом современных представлений морфо-функциональной организации ГАМК-ергической нейромедиаторной системы, рассматриваются патогенетические особенности токсического действия физиологически активных веществ, нарушающих её функционирование. Обсуждаются принципы терапии острых отравлений ГАМК-антагонистами. Пособие предназначено для студентов Факультетов военного обучения, проходящих обучение по программе кафедры «Медицинской профилактики и защиты».
Рекомендуется Учебно-методическим объединением по медицинскому и фармацевтическому образованию вузов России в качестве учебного пособия для студентов факультетов военного обучения медицинских ВУЗов.
СОДЕРЖАНИЕ Введение Принятые сокращения Глава 1 ОБЩИЕ ВОПРОСЫ НЕРВНОЙ РЕГУЛЯЦИИ 1.1.Межклеточные взаимодействия – основа регуляции 1.2.Особенности нервной регуляции Глава 2 ГАММА – АМИНОМАСЛЯНАЯ КИСЛОТА – МЕДИАТОР ТОРМОЖЕНИЯ 2.1. Роль и место ГАМК в организме 2.2. Этапы метаболизма ГАМК 2.3. Биосинтез и катаболизм ГАМК 2.4. Депонирование, экзоцитоз и обратный захват ГАМК 2.5. ГАМК - зависимая мембрана и рецепция ГАМК 2.6. Классификация ГАМК - рецепторов 2.6.1. Структурно-функциональные особенности ГАМКА – рецепторов 2.6.2. Структурно-функциональные особенности ГАМКС – рецепторов 2.6.3. Структурно-функциональные особенности ГАМКВ – рецепторов 2.7. Виды ГАМК – ергического торможения Глава 3 ФИЗИОЛОГИЧЕСКИ АКТИВНЫЕ ВЕЩЕСТВА, НАРУШАЮЩИЕ ФУНКЦИОНИРОВАНИЕ ГАМК - ЕРГИЧЕСКОЙ НЕЙРОМЕДИАТОРНОЙ СИСТЕМЫ 3.1. Физиологически активные вещества, нарушающие синтез и депонирование ГАМК 3.2 Физиологически активные вещества, нарушающие процессы экзоцитоза ГАМК 3.3 Физиологически активные вещества, нарушающие процессы обратного захвата и катаболизма ГАМК 3.4 Физиологически активные вещества, нарушающие процессы рецепции ГАМК 3.4.1. Конкурентные эффекторы ГАМК 3.4.2. Неконкурентные эффекторы ГАМК Глава 4 Заключение Терминологический словарь Рекомендуемая литература
ВВЕДЕНИЕ
Цель данного раздела курса - ознакомить студентов с вопросами токсикологии конвульсантов, опосредующих свою активность посредством нарушения функционирования ГАМК - ергической нейромедиаторной системы. Известно огромное количество химических соединений природного и синтетического происхождения, способных вызывать развитие судорожного синдрома у человека и экспериментальных животных. В одних случаях судорожная активность является неспецифическим проявлением токсического действия. В других, - токсиканты действуют селективно. Последние относятся к физиологически активным веществам (ФАВ) судорожного действия. При отравлении конвульсантами могут возникать клонические (коразол, цикутотоксин), клонико-тонические (физостигмин, фосфороорганические яды) и тонические (стрихнин) судороги.
Несмотря на многообразие механизмов, посредством которых нейротропные ФАВ вызывают судорожную реакцию, все они могут быть отнесены либо к гиперактивации процессов возбуждения, либо угнетению процесов торможения. Тормозные процессы в нервной системе осуществляются посредством пресинаптических и постсинаптических механизмов. В обоих случаях реализуется действие специфических нейромедиаторных систем мозга. Основным нейромедиатором торможения является g - аминомаслянная кислота (ГАМК). Идентифицированная около пятидесяти лет тому назад как нейромедиатор, ГАМК привлекает к себе пристальное внимание исследователей в связи с исключительно важной ролью в осуществлении интегративных функций нервной системы. Использование современных методов исследований и новейших технологий позволило в последние годы достичь определенных успехов в понимании морфо-функциональных особенностей ГАМК – ергической нейромедиаторной системы. В то же время, обобщённая структурно-функциональная «синаптическая модель мозга», казалось бы, логично выстроенная, все еще далека от совершенства в плане описания тормозных процессов ЦНС. В связи с этим, одной из кардинальных задач современной нейробиологии является детальное выяснение механизмов изменений синаптической передачи в тормозных нейронах в условиях различного функционирования организма. С этой целью активно проводятся исследования как субъединичного состава молекулярных рецепторов ГАМК, так и биохимических каскадов, запускаемых при их модуляции. ФАВ, модулирующие активность ГАМК – ергической нейромедиаторной системы, незаменимы при изучении структуры и функции ГАМК-ергической нейротрансмиссии. Обладая широким спектром физиологической активности, они широко используются в качестве «фармакологических зондов». Их использование в научных исследованиях обеспечило определённый прогресс в нейрофармакологии анксиолитиков, снотворных, противосудорожных средств и других фармакологических препаратов. Понимание механизмов функционирования, знание «уязвимых» участков ГАМК-ергической нейропередачи является отправной точкой для уточнения патогенеза различных заболеваний, выявления новых ФАВ (как лекарственных препаратов, так и токсикантов) с заданными свойствами. Вопросы метаболизма ГАМК и возможности её антиметаболизма составляют содержание настоящего учебного пособия. Знание этих вопросов является необходимой научной базой для подготовки квалифицированных специалистов лечебно-профилактического профиля.
ПРИНЯТЫЕ СОКРАЩЕНИЯ АДЦ - аденилатциклаза АМФ - аденозинмонофосфат АОУК - аминооксиуксусная кисота АРГ - аргинин АТФ - аденозинтрифосфат БЗД - 5-арилбенз-1,4-диазепины БЦГ - бициклогептан (норборнан) - БЦО - бициклические эфиры ортокарбоновых кислот БЦОБ - бициклоортобензоат БЦОК - бициклоортокарбоксилат БЦФ - бициклические эфиры кислот фосфора (4-алкил-БЦФ) ВЗМО - высшая занятая молекулярная орбиталь ВПСП - возбудимый постсинаптический потенциал
ГАМК (GABA) - гамма-аминомасляная кислота ГАМКа(в,с) - подтипы рецепторов ГАМК ГАМКа РКК - ГАМК-рецепторно-канальный комплекс (ГАМК-бензодиазепин-хлор - ионный комплекс) ГАМК-Т - ГАМК-трансфераза ГДК (GAD) - глутаматдекарбоксилаза ГЛИ - глицин ГЛУ - глутаминовая кислота ГМФ - гуанозинмонофосфат ГОМК - гамма-оксимасляная кислота ГТФ - гуанозинтрифосфат ГУЦ - гуанилатциклаза ГЭБ - гематоэнцефалический барьер ДВС - синдром диссеминированного внутрисосудистого свёртывания крови ДСТА - дисульфатетраазаадамантан КРБ - b-карболин ЛИЗ - лизин М - медиатор МАО - моноаминооксидаза Мб - метаболит МСП - модулятор структурной перестройки НСМО - низшая свободная молекулярная орбиталь ПКТ - пикротоксинин ПЛ - пиридоксаль ПЛК - пиридоксаль-киназа ПЛФ - пиридоксальфосфат Р - рецептор ССАД - сукцинатсемиальдегид ТПСП - тормозной постсинаптический потенциал ФАВ - физиологически активное вещество Ф - фермент ФДЭ - фосфодиэстераза ХИК - хлор-ионный канал ХИН - хлор-ионный насос ХИФ - хлор-ионофор ц-АМФ - цикло-аденозинмонофосфат ц-ГМФ - цикло-гуанозинмонофосфат ЦИС - цистеин ЦНС - центральная нервная система Э - эффектор ЭЭГ - электроэнцефалография САСА - цис-аминокротоновая кислота DE - эффективная доза DC - конвульсивная (судорожная) доза DL - летальная (смертельная доза) GAT - электрогенные транспортёры ГАМК, осуществляющие её реаптейк ТРMPA - 1,2,5,6- тетрагидропиридин-4-метилфосфиновая кислота TBOB - третбутилбициклоортобензоат TBPO - третбутилбициклофосфат TBPS - третбутилбициклофосфоротионат g - ГАМК gЛ - ГАМК-литик gМ - ГАМК-миметик gР - ГАМК-рецептор ОБЩИЕ ВОПРОСЫ НЕРВНОЙ РЕГУЛЯЦИИ
ГАМК создает физиологически адекватный фон для нейрохимической организации регуляторных процессов как центральной, так и периферической нервной системы. Функционирование гомеостатических механизмов на организменном уровне тесно связано с поддержанием постоянства концентрации ГАМК в тканях мозга, которое обуславливает сбалансированность процессов возбуждения и торможения в нервных клетках. Снижение активности ГАМК-ергических механизмов сопровождается развитием дисстресса, инсомнии, тревожно-фобических и агрессивных реакций, судорожных состояний; приводит к нарушению ряда когнитивных функций. Повышение активности ГАМК-эргическрой системы провоцирует развитие депрессии, шизофрении, паркинсонизма. Экспериментальные и клинические исследования последних лет позволяют полагать, что формирование (и развитие) той или иной патологии обусловлено как нарушениями центральных ГАМК-ергических механизмов, так и нарушениями ГАМК-ергической нейротрансмиссии периферической нервной системы. Более того, ГАМК выступает не только как нейротрансмиттер, но и как паракринный эффектор, регулирующий различные физиологические механизмы на периферии.
Несомненно, что метаболизм ГАМК имеет первостепенное значение для жизнедеятельности. Нарушения метаболизма ГАМК опасны для организма в связи с возможными извращениями процессов регуляции в тканях головного мозга и последующим искажением нормальных физиологических реакций организма.
Этапы метаболизма ГАМК. Малоспецифичный для тканей мозга биосинтез ГАМК иногда реализуется в клетках отдельных тканей в связи с функционированием ряда аминотрансфераз, контролирующих переаминирование при катаболизме различных a-аминокислот и a-кетокислот, например:
В частности, к числу таких малоспецифичных процессов относится и переаминирование с участием глутаминовой (ГЛУ) и кетоглутаровой кислот, а также различных полифункциональных альдегидов, в том числе гамма-оксомасляной кислоты (янтарного полуальдегида). И именно в этом последнем случае имеет место равновесное образование ГАМК:
Такое равновесное образование ГАМК, реализующееся под контролем малоспецифичной ГАМК-трансаминазы, получило название «ГАМК-шунт» (см.2.3). Естественно, что равновесие «ГАМК-шунта» будет сдвинуто в ту или иную сторону в зависимости от избытка какого-либо из реагентов этой системы. Однако, оказалось, что ни один из четырех реагентов системы "ГАМК-шунт" (в том числе ГЛУ и ГАМК) не способен преодолевать гематоэнцефалический барьер (ГЭБ), в связи с чем их экзогенное вмешательство в нейрональный метаболизм ГАМК (при отсутствии патологии) оказывается практически невозможным. Однако в условиях церебральной патологии реагенты. «ГАМК-шунта» могут преодолевать ГБЭ; в частности, ГАМК способна проникать в верхние отделы промежуточного мозга. В связи с этим, при некоторых сосудистых заболеваниях головного мозга (сопровождающихся нарушением внимания, памяти, речи), при систематических головокружениях и головных болях, а также после инсульта и некоторых травм мозга рекомендовано экзогенное введение ГАМК. Начиная с 70-х годов, ГАМК стала использоваться в терапевтических и профилактических целях; коммерческие названия разных фармакопейных форм ГАМК: гаммалон, ганеврин, аминалон. Биосинтез и катаболизм ГАМК
В нервной ткани ГАМК образуется путем декарбоксилирования глютамата под влиянием энзима (КФ, 4.1.1.15), чаще называемого L-глутамат- 1-декарбоксилазой (ГДК;GAD) или декарбоксилазой глутаминовой кислоты (ДГК). ГДК является протеидом гексамерного типа с молекулярной массой около 90000 и оптимумом рН=7,0. Кофермент ГДК строго доказан - им является пиридоксаль-5-фосфат (ПЛФ), который поступает в ЦНС с кровотоком, а в организм вообще - в качестве основного компонента витамина В6:
Кофермент обратимо связан с апоферментом (полипептидным фрагментом) за счет конденсации альдегидной группы ПЛФ с w-аминогруппой лизинового фрагмента полипептида (ЛИЗ). Одновременно с этим имеет место и дополнительное связывание кофермента и апофермента за счет электростатического взаимодействия фосфатной группировки ПЛФ с аргининовыми фрагментами полипептида (АРГ), а также (предположительно) фенольного гидроксила ПЛФ с атомом тяжелого металла (главным образом Fе+++), ковалентно связанного с меркаптогруппами цистеиновых фрагментов полипептида (ЦИС).
ГДК характеризуется исключительной субстратной специфичностью: фермент in vivo контролирует декарбоксилирование только L-ГЛУ и репрессируется только самим метаболитом, то есть ГАМК. Репрессорная способность ГАМК, проявляется посредством использования «стандартного» механизма гормональной регуляции. То есть, если ГДК рассматривать как внутренний структурный элемент какого-то определенного участка мембраны синаптосомы; внешним элементом этого же участка мембраны является синапторецептор ГАМК (gРС).
При избытке ГАМК в нейрональной среде имеет место ее равновесная рецепция на ГАМК-зависимых синапторецепторах, сопровождающаяся структурной перестройкой этого участка мембраны. В результате такой перестройки из разобщенных элементов мембраны формируется единый структурный блок ГАМК-[синапторецептор ГАМК]. Глутаминовая кислота является основным источником синтеза ГАМК в тканях мозга; синтез ГАМК из путресцина, спермидина и пирролидона имеет второстепенное значение. Её концентрация в мозговой ткани достигает 10 мкМ/г ткани. Предполагается, что около 8-10% глутамата может превращатьтся по альтернативному (по отношению к его участию в цикле трикарбоновых кислот) пути с образованием ГАМК. Данные, характеризующие локализацию ГДК (обнаружена только в нейронах, высвобождающих ГАМК), свидетельствуют о том, что основная функция ГАМК-шунта не связана с метаболизмом глюкозы. Источником веществ-предшественников глютамата является цикл Кребса (рис.2.2).
Рис.2.2. Образование и окисление глутамата в головном мозге. 1 – глутаматдегидрогеназа, 2 – аспартатаминотрансфераза, 3 – аланинаминотрансфераза, 4 - тирозинаминотрансфераза, 5 – трансаминаза ГАМК.
ГДК является ключевым ферментом, лимитирующий скорость синтенза ГАМК. Существуют две основные изоформы глутаматдекарбоксилазы: GAD67 и GAD65. GAD67 распределена во всей цитоплазме нейронов, a GAD65 в основном локализована в пресинаптических терминалях ГАМК-ергических интернейронов. Экспрессия последнего фермента существенно меняется в зависимости от уровня активности нейрона, что указывает на его особую значимость в регуляции ГАМК-ергической передачи. GAD67 имеет высокое сродство к пиридоксальфосфату, и поэтому может быть активированной постоянно. GAD65 имеет низкое сродство к пиридоксальфосфату и её активность регулируется доступностью кофактора. Характерно, что мутантные мыши, (у которых отсутствует GAD65) при нормальном уровне ГАМК, более чувствительны к судорогам. Катаболизм ГАМК осуществляется митохондриальным энзимом К.Ф,2.6.1.19, иначе называемым ГАМК-трансаминазой (ГАМК-Т), превращающим аминокислоту в сукцинатсемиальдегид (ССАД), который в свою очередь, окисляется дегидрогеназой сукцинатсемиальдегида до янтарной кислоты с последующей утилизацией в цикле Кребса (рис.2.3.).
Рис.2.3. Схема обмена ГАМК в ЦНС ЦТК - цикл трикарбоновых кислот; ГК - глутаминовая кислота; ДГК - декарбоксилаза глутаминовой кислоты; ГАМК-Т - ГАМК-трансаминаза; ССАД – сукцинатсемиальдегид
Синтез и депонирование ГАМК происходят, по-видимому, в разных компартментах клетки. Шунт ГАМК представляет собой мостик между двумя этими компартментами, в одном из которых (в нервных окончаниях) ГАМК синтезируется, а после выделения деградирует в компартментах глиальных клеток.
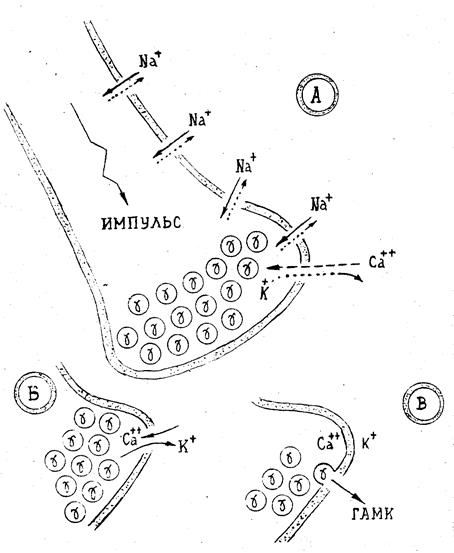
Высвобождение ГАМК в синаптическую щель протекает в соответствии с общими закономерностями высвобождения нейромедиаторов: экзоцитоз реализуется когда распространяющийся внутринейронно биохемоэлектрический импульс достигает окончания нейрона, где размещены синаптические везикулы с депонированной в них ГАМК. Непосредственно предшествующим экзоцитозу актом является формирование неспецифического кальций-калий-ионного канала. ГАМК при этом выполняет функцию модулятора экзоцитоза: когда её концентрация в синаптическом пространстве достигает определённого «критического» уровня, включаются механизмы прекращения экзоцитоза и одновременной трансформации кальций-калиевого канала в кальций-калиевый насос.
Рис. 2.4. Принципиальная схема экзоцитоза ГАМК (g): А. Функционирование ионных каналов. Б. Условия экзоцитоза. В. Одна из "сцен" экзоцитоза.
Экзоцитоз фактически начинается с момента достижения биохемоэлектрическим импульсом пресинаптической мембраны и формирования в ней кальций-калий-ионного канала (рис.2.4.). Сквозь этот канал по закону пассивной диффузии (см. 1.1) в нейрональную среду начинает поступать поток ионов Са++, а из нейрона в синаптическую щель — регенеративный поток ионов К+; при этом, градиент ионов Na+ остается неизменным. По мере увеличения в нейрональной среде концентрации ионов Са++, а в синаптическом пространстве концентрации ионов К+ при остром дефиците в окончании нейрона ионов Nа+ экзоцитоз ускоряется. Скорость экзоцитоза достигает максимума, когда концентрации ионов составляют: [K+]синапс=0,04-0,05 эВ и [Са++]нейрон=0,0009-0,001 эВ. Когда же концентрация ионов Са++ достигает уровня 0,001-0,005 эВ, то экзоцитоз ГАМК частично тормозится. При превышении же уровня 0,01-0,015 эВ Са++—экзоциитоз прекращается полностью. Этот эффект объясняется реализацией принципа обратной связи: ГАМК (как и многие другие медиаторы) выполняет функции модулятора экзоцитоза. Это означает, что, когда концентрация ГАМК в синаптическом пространстве достигает определенного («критического») уровня, то имеет место разновесная рецепция медиатора на пресинаптической мембране, благодаря чему «включаются» механизмы, приводящие к прекращению экзоцитоза ГАМК и одновременно к трансформации кальций-калий-ионного канала в соответствующий насос. Такой насос обеспечивает метаболический перенос ионов кальция и калия в обратных направлениях (Са++ — из нейрона, К+ — в нейрон) с восстановлением исходных градиентов концентраций этих ионов. Роль ионов кальция заключается в стимуляции процессов фосфорилирования белков, а следовательно- в последующем слиянии мембран синаптических пузырьков с пресинаптической мембраной. Эти эффекты кальция опосредованы особым белком- синапсином I, локализованным на поверхности везикул. Весь цикл рассмотренных эффектов завершается за весьма короткое время около 0,03 с. и затем - все механизмы пресинаптической мембраны вновь готовы к следующему акту экзоцитоза. Следует отметить, что высвобождение ГАМК возможно как в виде мультимолекулярных квантов (что происходит при слиянии синаптических везикул с плазматичесчкой мембраной), так и в виде постоянной неквантовой утечки медиатора из окончания аксонов в состоянии покоя (см. 2.7). После высвобождения ГАМК в синаптическую щель, помимо взаимодействия нейромедиатора с соответствующим рецептором, происходит ее захват электрогенными транспортерами. Транспортёры, которые переносят нейромедиаторы в нервное окончание, отличаются от тех, которые транспортируют медиатор в синаптические везикулы. В настоящее время идентифицировано три таких транспортера (GAT1, GAT2 и GAT3), локализующихся как в астроцитах, так и в самих нейронах. Эти транспортеры различаются спецификой клеточной и региональной локализации. Так, например, GAT1 и GAT3 широко представлены в различных отделах мозга (показательно, что фармакологический профиль глиальных и нейрональных транспортёров ГАМК различен); GAT2 представлен в периферических тканях: сердце, печень, почки и т. д. Поскольку транспорт является электрогенным, анион аминокислоты переносится вместе с двумя катионами натрия и одним анионом хлора. Большая часть выделившейся в синаптическую щель ГАМК удаляется оттуда посредством высокоафинного Na+-зависимого активного захвата как нервными окончаниями, так и клетками глии. Каждая функциональная группа транспортёра связана с определённой белковой молекулой (или с их семейством). Большинство из них имеют до 12 трансмембранных сегментов (доменов) и образуют канало-подобные структуры. Для «работы» транспортёров необходимы внеклеточные ионы натрия и хлора, которые транспортируются одновременно с медиатором.
Считается, что быстрые тормозные постсинаптические токи (ТПСТ) опосредованны ионотропными рецепторами ГАМК, расположенными непосредственно в пределах активной зоны; медленное тоническое ГАМК-опосредованное торможение обеспечивается внесинаптическими ионотропными рецепторами ГАМК. Фазное торможение нейронов определяется дискретным выбросом в синаптичекую щель такого количества ГАМК, которое создаёт высокие концентрации нейромедиатора и эффективно воздействует на постсинаптические ГАМК-рецепторы. Тоническое торможение связано с постоянной слабой активацией ГАМК-рецепторов. Роль тонического ГАМК-опосредованного тока состоит в поддержании определенного значения потенциала на мембране и соответствующей модуляции возбудимости клетки. Другой функцией тонического торможения, вероятно, является шунтирование фазных пре- и постсинаптических трансмембранных токов при развитии потенциалов действия. Шунтирование быстрых токов происходит из-за того, что тоническая проводимость, связанная с наличием открытых каналов ГАМК-рецепторов, снижает сопротивление мембраны. Повышение электрической проводимости (уменьшение сопротивления) мембраны приводит к падению амплитуды потенциала действия, поступающего в пресинаптический участок аксона, уменьшая тем самым вход Са2+ и снижая вероятность выброса медиатора. Механизмы тонического ГАМК-ергического торможения изучены недостаточно полно. Согласно одной из гипотез, постоянная составляющая ГАМК-ергического тока представляет собой суммацию спонтанных ТПСТ, возникающих в ответ на спонтанный, не индуцированный пресинаптическим импульсом, выброс ГАМК. Согласно другой, - тонический ГАМК-ергический ток возникает за счет диффузии ГАМК во внесинаптическое пространство (в результате «перелива») и последующей активации внесинаптических рецепторов этого трансмиттера, свойства которых отличны от свойств синаптических рецепторов. Функция внесинаптических ГАМК-рецепторов, по всей видимости, заключается в детектировании внеклеточной концентрации ГАМК и поддержании соответствующего уровня тонического торможения (рис.2.6.). Помимо молекул ГАМК, покидающих синаптическую щель, определенную роль в повышении внеклеточной концентрации ГАМК (следовательно, и в тоническом ГАМК-опосредованном торможении) может играть функционирование ГАМК-транспортеров в обратном направлении, высвобождение ГАМК астроцитами и снижение активности ГАМК-транаминазы. Существует точка зрения, что происхождение ГАМК-ергического тонического тока связано со спонтанным (без участия нейротрансмиттера) открытием каналов ГАМК-рецепторов. Если этот (без предварительного высвобождения агониста и воздействия на рецептор) процесс действительно может происходить, то конкурентные антагонисты ГАМК (например, SR95531) не должны оказывать влияния на тонический ток, а неконкурентные блокаторы ГАМК-рецепторов (такие, как пикротоксин) будут подавлять тонический ток. Кроме того, в рамках рассматриваемой гипотезы повышение внеклеточной концентрации ГАМК не должно усиливать тонического торможения. Не исключена возможность существования двух компонентов тонического тока: зависящего от внеклеточной концентрации ГАМК и не зависящего, (опосредованного спонтанным открыванием ГАМК-ергических каналов). Вышеописанные типы торможения определенным образом задействованы в процессах эпилептогенеза. Так при моделировании эпилепсии на животных фиксируется гибель только определённых видов интернейронов; отмечается снижение интенсивности дендритного, но не соматического торможения. Аппликация каината повышает эффективность ГАМК-ергического торможения в интернейронах, но снижает интенсивность такого влияния на пирамидные клетки. Важно отметить, что все случаи снижения ритма биосинтеза ГАМК. приводящие к постепенному истощению запаса этого медиатора, и как следствие. - к последующим извращениям нервной регуляции, характеризуются наличием скрытого периода. Депрессия, возникающая при отравлениях сублетальными дозами рассматриваемых ФАВ, оказывается относительно устойчивой - продолжается десятки часов. Противоположный феномен - ускорение нормального ритма биосинтеза ГАМК в принципе невозможен, так как производительность биосинтеза в норме строго лимитирована количеством регулирующего этот процесс фермента (ГДК). Однако возрастание уровня ГАМК в нейроне иногда наблюдается вследствие увеличения в тканях ЦНС производительности «ГАМК-шунта», при введении экзогенного глутамина. Однако этот феномен не сопровождается существенным нарушением нервной регуляции по той причине, что при достижении в нейрональной среде определённого уровня ГАМК реализуется его способность к репрессии активности ГДК. К тому же, как только содержание ГАМК в нейрональной среде достигает критического уровня, имеет место депонирование нейромедиатора в синаптические везикулы. Возможность экзогенного влияния на характер и механизм депонирования ГАМК до конца не изучен (см.2.4). Пока неизвестно ни одного соединения, которое бы in vivo выступало в роли специфичного активатора или ингибитора депонирования ГАМК. Известно лишь, что депонирование ускоряется в присутствии ионов хлора, достигая максимума при концентрации 0.05 М С1-. Кроме того, опытами in vitro показано, что депонирование ГАМК тормозится резерпином и некоторыми производными фенотиазина и дибензазенина. в частности, аминазином и имипрамином.
Диагностика столбняка основана на фиксации одновременно нескольких клинических признаков (обильное потоотделение, тризм, сардоническая улыбка, нисходящее распространение ригидности, рефлекторная возбудимость и гипертермия) и результатах иммунохимической реакции крови. Идентификация тетаноспазмина в различных пробах проводится иммунными методами; неспецифическая индикация возможна посредством классических аналитических реакций на белки. Дезактивация тетаноспазмина осуществляется с использованием щелочей, растворов веществ окислительно-хлорирующего действия, высоких температур (при 80-100°С). Лечение столбняка - комплексное (симптоматическая терапия сочетается с Высокая летальность, трудности ранней диагностики и сложность организации системы мероприятий по защите и противодействию, обусловливает возможность использования экзотоксинов столбняка в качестве смертоносных химических агентов, например, в средствах микстовых поражений. Таким образом, предполагается возможность взаимодействия блокаторов ХИК как со специфическим участком связывания внутри канала с последующей его закупоркой, так и с участком связывания вблизи канала и дальнейшими конформационными изменениями постсинаптической мембраны, сопровождающимися нарушением кинетики ионофора. Считается, что даже для сходных по химической структуре соединений имеет место гетерогенность участков связывания. В обзорах, обобщающих данные по биологической активности непрямых ГАМК-антагонистов (И.В. Мартынов, В.И. Фетисов, В.Б. Соколов «Бициклические ортоэфиры кислот фосфора» 1989; А.И. Головко, С.И. Головко. С.Ю Зефиров, Г.А. Софронов «Токсикология ГАМК-литиков»1996) подчеркивается, что наиболее токсичными синтетическими блокаторами хлорионофоров являются трет-бутил-БЦФ-ат (DL.50 б.м.= 0,038 мг/кг); трет-бутил-цианофенил (DL50 б.м.=0,06 мг/кг) и экзо-цис-5,6-дихлор,-2,2-дициано-3.3 -бис(трифторметил), иначе называемый «бициклогептан» или «норборнан» (DL50собаки=0,045 мг/кг). Бициклогептан (БЦГ) в наномолярных концентрациях способен угнетать стимулируемый ГАМК захват радиоактивного хлора и тормозить специфическое связывание 35S-TBPS и ТВОВ; по степени аффинности к белку ионофора значительно превосходит трет-БЦФ-ат, ДСТА, пикротоксин (IC.50 соответственно 46.72.700 и 5000 нМ). Токсическое действие данных соединений проявляется клонико-тоническими судорогами в экспериментах на различных видах ЛЖ. Для бициклофосфатов и бициклоортобензоатов характерно стремительное развитие интоксикации и гибель в течение 1-30 минут, для БЦГ, - отсроченное развитие симптоматики и гибель в течение нескольких суток. Отличаясь высоким сродством к специфическому участку связывания хлорионофора, БЦГ вызывает развитие судорожных пароксизмов при блокаде не менее половины хлор-ионных каналов ЦНС. При этом отмечается преимущественное ингибирование ХИК ГАМКА-РКК в стриатуме и мозжечке, в то время как во фронтальной коре плотность хлор-ионофоров остается в пределах исходных показателей. Профилактика и купирование судорожного синдрома «конвульсантами клеточной структуры» связаны с определенными трудностями (Головко А.И., Головко С.И. с соавт. 1996). Проведенные биохимические и радиолигандные исследования БЦФ-атов и БЦГ выявили общность их механизма действия, что позволяет отнести их к непрямым антагонистам ГАМКА-РКК. Данные соединения, так же как и пикротоксинин, линдан, в период судорожной активности вызывают значительное снижение уровня ц-АМФ и увеличение уровня ц-ГМФ в мозжечке. В то же время. БЦГ, в отличие от БЦФ-атов, ДСТА. пикротоксина, является необратимым селективным ингибитором ХИК ГАМКА-РКК. Наряду с БЦГ. необратимыми ингибиторами ХИК ГАМКА-РКК являются также пара- и меча- изотиоцианатные аналоги третбутилбициклоортобензоата и фотоафинные лиганды хлорионофора триоксабициклооктановой структуры
Клиническая картина проявления судорожного симптомокомплекса в условиях хронического отравления блокаторами ГАМК-зависимых хлор-ионных каналов характеризуется более выраженным психо-вегетативным сопровождением и стёртостью фазности течения интоксикации. Дисбаланс тормозных и возбуждающих процессов, нарушение интегративной деятельности мозговых структур вследствие нарушения ГАМК-ергических механизмов тормозного контроля в стрианигропаллидарной системе и формирование генератора патологически усиленного возбуждения в стриатум определяет избирательный физиологический эффект. Проявлением неспецифических защитных механизмов является токсический стресс, протекающий в условиях нарушения модуляторного влияния ГАМК на сегментарную и рассогласования функционирования эрготропных и трофотропных отделов надсегментарного аппарата вегетативной нервной системы. Лидирующим патогенетическим механизмом является гипоксия. В результате реализации специфических и неспецифических патогенетических механизмов, в результате подавления активности физиологических систем мозга и соответствующих механизмов гомеостаза происходит стабилизация патологического процесса, то есть формирование устойчивого патологического состояния. Совокупность кардиореспираторных нарушений и проявлений судорожного синдрома, замыкающихся в «порочный круг», формирует типовые патологические процессы (гипоксию, шок, кому), приводящие к гибели. Перевод на более низкий информационно-энергетический уровень функционирования ЦНС, в ряде случаев, обеспечивает мобилизацию энергетических и пластических резервов нервной системы и определяет сохранение жизни. Углубление представлений относительно функционирования ГАМК-ергической нейромедиаторной системы, достигнутое в последние годы, позволяет ставить вопрос о возможных путях модуляции ГАМК-ергических синаптических процессов посредством использования не только традиционных эффекторов ГАМК-рецепторно-канального комплекса (барбитураты, бензодиазепины), но и разрабатываемх селективных агонистов ГАМК, воздействующих на определённые подтипы субъединиц ГАМК –БД- рецепторно-канального комплекса (см.2.6.). В последнем случае повышается противосудорожная активность и уменьшается вероятность развития побочных эффектов, связанных с седатацией, миорелаксаций, когнитивными нарушениями тех же бензодиазепинов. Определённые надежды связывают и с направлением экспериментальной терапии, ориентирующимся на увеличение концентрации ГАМК в синаптической щели посредством блокады её обратного захвата и ингибирования активности ГАМК-трансаминазы. Что касается полипрагмазии, то здесь необходимо учитывать, что совместное действие средств специфической терапии характеризуется суммацией и взаимопотенцированием как терапевтического, так и побочного эффектов. И, в этой связи, принципиально важным является введение в схему купирования судорожных состояний препаратов, воздействующих на неспецифические звенья патогенеза: уменьшение про
|
||||||||||||||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 799; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.142.135.24 (0.019 с.) |