Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Депонирование, экзоцитоз и обратный захват ГАМК.Содержание книги
Поиск на нашем сайте
В пресинаптических окончаниях ГАМК депонируется в везикулах (аксон ГАМК-реактивного нейрона содержит около 1000 везикул, в каждой из которых содержится более 10000 молекул ГАМК). Депо медиатора составляет приблизительно 10 млн. молекул/нейроне. Везикуляция ГАМК осуществляется с помощью специальных транспортеров, которые используют протонный градиент, создаваемый везикулярными АТФ-азами. Показательно то, что механизмы везикулярного захвата возбуждающего или тормозного нейротрансмиттера (глутамата и ГАМК соответственно) принципиально различны. Так анионы глутамата непосредственно перемещаются в везикулы по градиенту концентрации Н+, которая в этих структурах высока. В случае ГАМК по протонному градиенту вначале движутся анионы хлора, которые затем обмениваются на анионы ГАМК. Белок, участвующий в процессе поглощения глутамата везикулами, идентифицирован S. Takamori, J. S., Rhec с сотрудникамим. (2000). Белок, опосредующий обмен хлора на ГАМК, до сих пор не определен. Известно, что захват ГАМК в везикулы происходит при вспомогательном участии фермента VGAT1, осуществляющего везикуляцию глицина, но не являющегося обязательным при упаковке ГАМК в везикулы. Тот факт, что эти два нейромедиатора имеют один и тот же везикулярный транспортёр, даёт основание полагать, что в некоторых нейронах ГАМК и глицин упаковываются вместе в одни и те же пузырьки. Высвобождение ГАМК в синаптическую щель протекает в соответствии с общими закономерностями высвобождения нейромедиаторов: экзоцитоз реализуется когда распространяющийся внутринейронно биохемоэлектрический импульс достигает окончания нейрона, где размещены синаптические везикулы с депонированной в них ГАМК. Непосредственно предшествующим экзоцитозу актом является формирование неспецифического кальций-калий-ионного канала. ГАМК при этом выполняет функцию модулятора экзоцитоза: когда её концентрация в синаптическом пространстве достигает определённого «критического» уровня, включаются механизмы прекращения экзоцитоза и одновременной трансформации кальций-калиевого канала в кальций-калиевый насос.
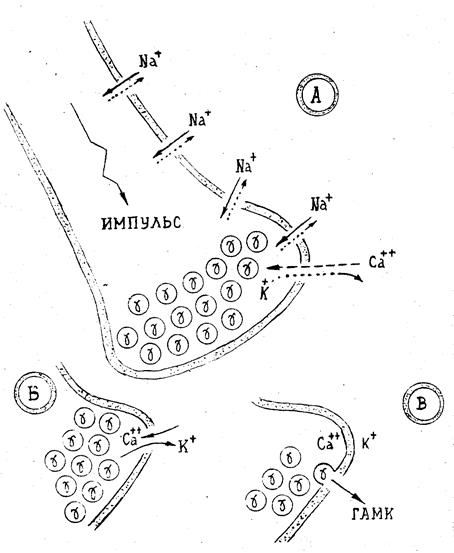
Рис. 2.4. Принципиальная схема экзоцитоза ГАМК (g): А. Функционирование ионных каналов. Б. Условия экзоцитоза. В. Одна из "сцен" экзоцитоза.
Экзоцитоз фактически начинается с момента достижения биохемоэлектрическим импульсом пресинаптической мембраны и формирования в ней кальций-калий-ионного канала (рис.2.4.). Сквозь этот канал по закону пассивной диффузии (см. 1.1) в нейрональную среду начинает поступать поток ионов Са++, а из нейрона в синаптическую щель — регенеративный поток ионов К+; при этом, градиент ионов Na+ остается неизменным. По мере увеличения в нейрональной среде концентрации ионов Са++, а в синаптическом пространстве концентрации ионов К+ при остром дефиците в окончании нейрона ионов Nа+ экзоцитоз ускоряется. Скорость экзоцитоза достигает максимума, когда концентрации ионов составляют: [K+]синапс=0,04-0,05 эВ и [Са++]нейрон=0,0009-0,001 эВ. Когда же концентрация ионов Са++ достигает уровня 0,001-0,005 эВ, то экзоцитоз ГАМК частично тормозится. При превышении же уровня 0,01-0,015 эВ Са++—экзоциитоз прекращается полностью. Этот эффект объясняется реализацией принципа обратной связи: ГАМК (как и многие другие медиаторы) выполняет функции модулятора экзоцитоза. Это означает, что, когда концентрация ГАМК в синаптическом пространстве достигает определенного («критического») уровня, то имеет место разновесная рецепция медиатора на пресинаптической мембране, благодаря чему «включаются» механизмы, приводящие к прекращению экзоцитоза ГАМК и одновременно к трансформации кальций-калий-ионного канала в соответствующий насос. Такой насос обеспечивает метаболический перенос ионов кальция и калия в обратных направлениях (Са++ — из нейрона, К+ — в нейрон) с восстановлением исходных градиентов концентраций этих ионов. Роль ионов кальция заключается в стимуляции процессов фосфорилирования белков, а следовательно- в последующем слиянии мембран синаптических пузырьков с пресинаптической мембраной. Эти эффекты кальция опосредованы особым белком- синапсином I, локализованным на поверхности везикул. Весь цикл рассмотренных эффектов завершается за весьма короткое время около 0,03 с. и затем - все механизмы пресинаптической мембраны вновь готовы к следующему акту экзоцитоза. Следует отметить, что высвобождение ГАМК возможно как в виде мультимолекулярных квантов (что происходит при слиянии синаптических везикул с плазматичесчкой мембраной), так и в виде постоянной неквантовой утечки медиатора из окончания аксонов в состоянии покоя (см. 2.7).
После высвобождения ГАМК в синаптическую щель, помимо взаимодействия нейромедиатора с соответствующим рецептором, происходит ее захват электрогенными транспортерами. Транспортёры, которые переносят нейромедиаторы в нервное окончание, отличаются от тех, которые транспортируют медиатор в синаптические везикулы. В настоящее время идентифицировано три таких транспортера (GAT1, GAT2 и GAT3), локализующихся как в астроцитах, так и в самих нейронах. Эти транспортеры различаются спецификой клеточной и региональной локализации. Так, например, GAT1 и GAT3 широко представлены в различных отделах мозга (показательно, что фармакологический профиль глиальных и нейрональных транспортёров ГАМК различен); GAT2 представлен в периферических тканях: сердце, печень, почки и т. д. Поскольку транспорт является электрогенным, анион аминокислоты переносится вместе с двумя катионами натрия и одним анионом хлора. Большая часть выделившейся в синаптическую щель ГАМК удаляется оттуда посредством высокоафинного Na+-зависимого активного захвата как нервными окончаниями, так и клетками глии. Каждая функциональная группа транспортёра связана с определённой белковой молекулой (или с их семейством). Большинство из них имеют до 12 трансмембранных сегментов (доменов) и образуют канало-подобные структуры. Для «работы» транспортёров необходимы внеклеточные ионы натрия и хлора, которые транспортируются одновременно с медиатором.
|
||||||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 289; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.15.198.149 (0.011 с.) |