Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Распад глюкозы в аэробных и анаэробных условияхСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
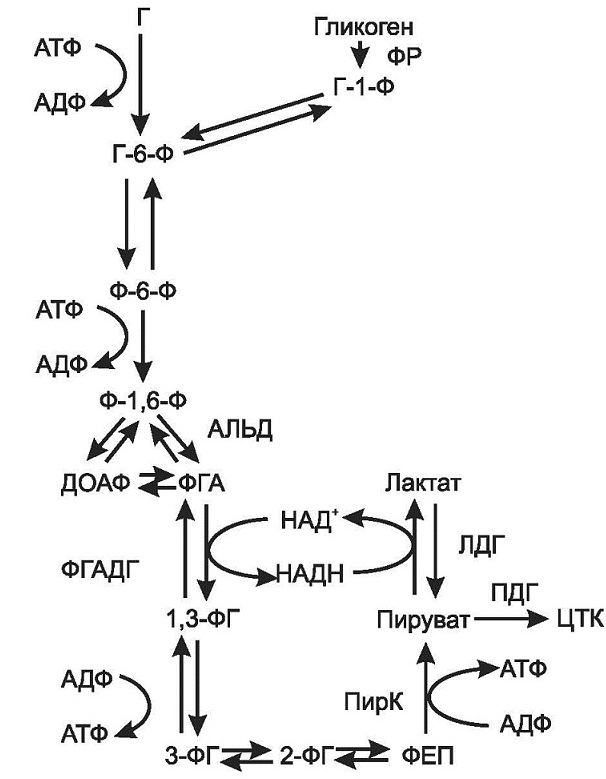
Катаболизм глюкозы – основной поставщик энергии для большинства клеток организма. Аэробный распад протекает в условиях достаточного снабжения тканей кислородом. Он включает несколько стадий: 1. Аэробный распад глюкозы до двух молекул пирувата в гиалоплазме клеток,который можно разделить на 2 этапа: а)подготовительный этап, в ходе которого глюкоза фосфорилируется и расщепляется на две молекулы фосфотриоз. б) основной этап, сопряженный с синтезом АТФ. В результате серии реакций фосфотриозы превращаются в пируват. 2. Превращение пирувата в ацетил-КоА и окисление последнего в цикле Кребса 3. Дыхательная цепь Глюкозо-6-фосфат, образованный в результате фосфорилирования глюкозы с участием АТФ, в ходе следующей реакции превращается в фруктозо-6-фосфат. Это обратимая реакция изомеризации протекает при участии глюкозофосфатизомеразы. Затем следует еще одна реакция фосфорилирования за счет АТФ, катализирует ее фосфофруктокиназа. В ходе этой реакции фруктозо-6-фосфат превращается в фруктозо-1,6-бисфосфат. Этот метаболит далее расщепляется на 2 триозофосфата: фосфоглицериноый альдегид (ФГА) и диоксиацетонфосфат (ДОАФ), реакцию катализирует альдолаза. В последующих реакциях гликолиза используется только ФГА, поэтому ДОАФ превращается с участием фермента триозофосфатизомеразы в ФГА. На этом подготовительный этап заканчивается. Основной этап аэробного распада включает реакции, связанные с синтезом АТФ. Сначала происходит окисление ФГАпри участии ФГА-дегидрогеназы. Этот фермент является НАД+-зависимымым. Восстановленный НАД+ затем окисляется в дыхательной цепи (перенос НАДН в митохондрии, где находится дыхательная цепь, происходит при участии челночных механизмов, это связано с тем, что митохондриальная мембрана непроницаема для НАДН). Кроме того, свободная реакция окисления этой реакции концентрируется в макроэргической связи продукта реакции. Продуктом реакции является 1,3-бисфосфоглицерат. Для фосфорилирования используется Н3РО4. В следующей реакции высокоэнергетический фосфат передается на АДФ с образованием АТФ. Катализирует реакцию фосфоглицераткиназа. АТФ в данной реакции образуется путем субстратного фосфорилирования. Продуктом реакции является 3-фосфоглицерат. В следующей реакции происходит перенос фосфата из 3 положения во второе с образованием 2-фосфоглицерата. Из него при отщеплении воды образуется фосфоенолпируват – макроэргическое соединение, фосфатная группа которого в следующей реакции переносится на АДФ с образованием АТФ при участии пируваткиназы (это вторая реакция субстратного фосфорилирования). Продуктом реакции является пируват. Во второй стадии пируват превращается в ацетил-КоА, который сгорает в цикле Кребса и дыхательной цепи (третья стадия) до СО2 и Н2О. Всего при аэробном окислении глюкозы выделяется 32 АТФ. Анаэробный гликолиз представляет собой распад глюкозы в анаэробных условиях (условиях недостаточного снабжения кислородом), поэтому он не зависит от работы митохондриальной дыхательной цепи. АТФ в гликолизе образуется путем субстратногофосфорилирования. Конечным продуктом является лактат (рис. 5). Все реакции анаэробного гликолиза (11 реакций) протекают в гиалоплазме. Первые 10 реакций до пирувата идентичны аэробному распаду. В последней реакции происходит восстановление пирувата в лактат цитозольным НАДН. Реакцию эту катализирует лактатдегидрогеназа. С помощью этой реакции обеспечивается регенерация НАД+ из НАДН без участия дыхательной цепи в ситуациях, связанных с недостаточным снабжением клеток кислородом. Роль акцептора водорода от НАДН (подобно кислороду в дыхательной цепи) выполняет пируват. Таким образом, значение реакции восстановления пирувата заключается не в образовании лактата, а в том, что данная цитозольная реакция обеспечивает регенерацию НАД+. К тому же лактат не является конечным продуктом метаболизма, удаляемым из организма (лактат – тупиковый метаболит). Он выводится в кровь и утилизируется: либо превращается в глюкозу (75%), либо при доступности кислорода окисляется до СО2 и Н2О (25%).
Рис.5. Гликолиз
Энергетический выход анаэробного гликолиза составляет 2 АТФ. Недостатками анаэробного гликолиза являются: малый энергетический выход по сравнению с аэробным распадом и накопление лактата, что приводит к ацидозу, усталости. Достоинства гликолиза: он полезен при выполнении кратковременной работы, является единственным источником энергии для эритроцитов (в них отсутствуют митохондрии), а также необходим в разных органах при дефиците кислорода (наложение жгута, падение давления, нарушение кровотока). Регуляция. Регуляторными ферментами гликолиза являются: гексокиназа, фосфофруктокиназа и пируваткиназа. Аллостерическими ингибиторами являются АТФ и цитрат. Стимулирует гликолиз инсулин, а тормозят глюкагон, глюкокортикостероиды. Катехоламины тормозят гликолиз в печени и активируют в мышцах.
ГЛЮКОНЕОГЕНЕЗ Некоторые ткани нуждаются в постоянном поступлении глюкозы. Когда поступление углеводов в составе пищи недостаточно, содержание глюкозы в крови некоторое время поддерживается в пределах нормы за счет расщепления гликогена в печени. Однако запасы гликогена в печени невелики, они практически полностью исчерпываются после суточного голодания. В этом случае в печени начинается глюконеогенез (ГНГ) – процесс синтеза глюкозы из веществ неуглеводной природы. Его основной функцией является поддержание уровня глюкозы в крови в период длительного голодания и интенсивных физических нагрузок. В первую очередь это важно для головного мозга (он не может обеспечивать потребности в энергии за счет окисления жирных кислот) и эритроцитов – в них аэробный путь распада веществ невозможен из-за отсутствия митохондрий. Процесс в основном (на 90%) протекает в печени и менее интенсивно в корковом веществе почек и слизистой тонкого кишечника. Субстратами ГНГ являются лактат, аминокислоты, глицерин. Аминокислоты включаются в ГНГ при длительном голодании или продолжительной мышечной работе. Глицерин освобождается в процессе липолиза из жировой ткани в период голодания или при длительной физической нагрузке. Лактат – продукт анаэробного гликолиза. Его источником в покое являются эритроциты, а при работе – мышцы. Таким образом, лактат используется в глюконеогенезе постоянно. Большинство реакций глюконеогенеза протекает за счет обратимых реакций гликолиза и катализируется теми же ферментами. Однако три реакции гликолиза необратимы. На этих стадиях реакции ГНГ протекают другими путями (обходные пути ГНГ). Первая необратимая реакция – образование из пирувата фосфоенолпирувата. В ГНГ это происходит в ходе двух реакций. Пируват в митохондриях карбоксилируется с образованием оксалоацетата при участии пируваткарбоксилазы – биотинзависимого фермента. Реакция протекает с использованием энергии АТФ. Дальнейшие превращения оксалоацетата протекают в цитозоле. Фосфоенолпируваткарбоксикиназа с затратой энергии ГТФ превращает оксалоацетат в фосфоенолпируват. Дальнейшие реакции ГНГ вплоть до образования фруктозо-1,6-бисфосфата протекают в цитозоле и катализируются гликолитическими ферментами. Затем следует еще одна необратимая реакция ГНГ, катализируемая фруктозо-1,6-бисфосфатазой, в которой происходит отщепление остатка фосфорной кислоты гидролитическим путем. Образующийся при этом фруктозо-6-фосфат гликолитическим ферментом изомеризуется до глюкозо-6-фосфата, от которого отщепляется остаток фосфорной кислоты при участии еще одного необратимо работающего фермента – глюкозо-6-фосфатазы. Образовавшаяся свободная глюкоза из клетки выходит в кровь. В ходе ГНГ расходуется 6 моль АТФ на синтез 1 моль глюкозы из пирувата или лактата. Ключевыми ферментами ГНГ, катализирующими необратимые обходные реакции, являются: пируваткарбоксилаза, фосфоенолпируваткарбоксикиназа, фруктозо-1,6-бисфосфатаза и глюкозо-6-фосфатаза. Активируют ГНГ аллостерически АТФ и цитрат, обязательным активатором первого фермента является ацетил-КоА. ГНГ усиливают гормоны: глюкагон, глюкокортикостероиды. Тормозит ГНГ гормон покоя и сытости – инсулин. Лактат, образовавшийся в ходе анаэробного гликолиза, не является конечным продуктом метаболизма. Использование лактата связано с его превращением в печени в пируват и с дальнейшим использованием последнего. Лактат, поступивший из интенсивно работающих мышц, выходит в кровь, а затем поступает в печень. В печени в лактатдегидрогеназной реакции лактат превращается в пируват, который включается в ГНГ, а образовавшаяся глюкоза поступает в кровь и поглощается скелетными мышцами. Эту последовательность событий называют «глюкозо-лактатным циклом», или «циклом Кори». Цикл Кори выполняет две важнейшие функции: обеспечивает утилизацию лактата и предотвращает лактатацидоз. Часть пирувата, образовавшегося из лактата (25%), окисляется печенью до СО2 и Н2О с освобождением энергии.
|
||||
|
|
Последнее изменение этой страницы: 2017-01-24; просмотров: 2588; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.133.127.131 (0.007 с.) |