Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Потенциал действия нервной клеткиСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
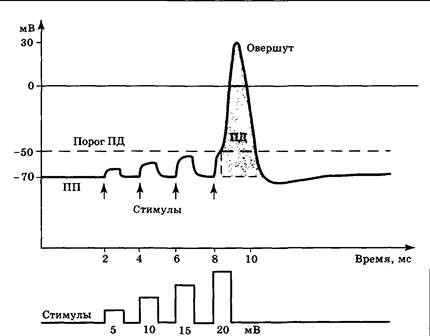
Мембрана нервной клетки постоянно находится в состоянии поляризации, и ее внутренняя сторона заряжена отрицательно по отношению к внешней. Процесс уменьшения разности потенциалов на мембране (снижение ПП по абсолютной величине) называется деполяризацией, а увеличение разности потенциалов (рост абсолютной величины ПП) — гиперполяризацией. Можно воздействовать на мембрану, используя стеклянный микроэлектрод. При подаче электрического тока и деполяризации мембраны на 10, 15, 20 мВ и более по отношению к уровню ПП (рис. 3.9) разность потенциалов на мембране достигнет примерно −50 мВ и будет наблюдаться феномен «взрывообразного» ее подъема и последующего спада. Это явление было названо потенциалом действия (ПД). В случае развития ПД ответ клетки на стимул значительно превышает амплитуду самого стимула. Феномен потенциала действия чрезвычайно важен для понимания общих принципов работы нервной системы. Именно в форме ПД осуществляется ответ нервных клеток на значимые стимулы, и в этой же форме сигналы распространяются по мембранам нейронов.
Средняя длительность ПД составляет около 1,5 мс. При этом его восходящая фаза (фаза деполяризации) несколько короче (около 0,5 мс) нисходящей фазы (фазы реполяризации). На вершине ПД разность потенциалов меняет свой знак и достигает примерно +30 мВ (овершут). После ПД могут развиваться более длительные стадии следовой гиперполяризации или деполяризации, имеющие обычно небольшую амплитуду. Форма и амплитуда ПД стабильны в каждом конкретном нейроне и не зависят от силы запускающего стимула, развитие ПД нейрона происходит по принципу «все или ничего». Уровень потенциала, при котором деполяризация мембраны приводит к запуску ПД, называется порогом ПД и чаще всего составляет −50 мВ. Разница между ПП и порогом ПД называется критическим уровнем деполяризации. Чем ниже (по абсолютной величине) ПП, тем меньше критический уровень деполяризации и выше возбудимость нейрона. Ионы, участвующие в генерации ПД, те же, что и в случае потенциала покоя — Na+ и К+. При развитии ПД натрий входит в нейрон, а калий выходит. Ионные каналы, через которые они движутся, относятся к отдельному классу — потенциал-зависимым (электрочувствительным) ионным каналам. Транспортные белки этого класса обладают способностью находиться в открытом либо закрытом состоянии в зависимости от разности потенциалов на мембране клетки. Обычно это связано с наличием заряженных фрагментов белковой молекулы — створок. Такие створки чаще всего перекрывают проход канала, если заряд внутри нейрона находится на уровне ПП, и открывают его при деполяризации мембраны.
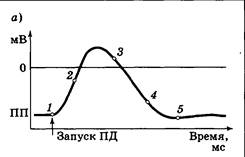
Если отдельно регистрировать движения (токи) ионов Na+ и К+ во время ПД, то можно получить следующие кривые (рис. 3.10). Натриевый ток возникает практически сразу после запускающего ПД стимула, в течение 0,3—0,4 мс достигает максимума, а затем снижается. Снижение и прекращение натриевого тока происходит на фоне еще не завершившегося ПД. Следовательно, время открытия Ка+-каналов ограничено, и их закрытие происходит вне зависимости от разности потенциалов на мембране. Калиевый ток, возникая позже натриевого, достигает максимума примерно через 1,0 мс после стимула. Противодействуя Na+-току, К+-ток выносит из клетки положительные заряды, возвращая разность потенциалов на мембране к уровню ПП. Таким образом, во время генерации ПД регистрируется более быстрый входящий ток ионов Na+ и более медленный выходящий ток ионов К+. Первый из них приводит к росту заряда внутри клетки (восходящая фаза ПД), второй — к возврату уровня потенциала покоя (нисходящая фаза ПД). Объяснить эти факты можно, если допустить существование у электрочувствительного Na+-канала не одной, а двух створок. Более «быстрая» из них открывает канал (активирует, А-створка); более «медленная» закрывает его (инактивирует, И-створка). В дальнейшем была доказана справедливость этой гипотезы, причем оказалось, что «медленная» створка прикреплена к внутренней стороне канала. В итоге деятельность Na+-канала в ходе генерации ПД можно представить следующим образом (рис. 3.11). На уровне ПП А-створка закрыта, И-створка открыта (состояние готовности, точка 1). При подаче стимула створки, не удерживаемые более отрицательным внутриклеточным зарядом, приходят в движение. А-створка очень быстро открывает проход внутри канала, и ионы Na+ начинают входить в нейрон; натриевый ток растет (точка 2). Скоро И-створка в своем движении достигает «верхнего положения»; каналы начинают закрываться, натриевый ток ослабевает и прекращается (точка 3).
Именно движение И-створки ограничивает время открытия канала, количество входящих в нейрон ионов Na+ и амплитуду ПД. В точке 3 Na+-канал пока еще не вернулся в начальную конфигурацию: хотя он и закрыт, створки его занимают положение, отличное от исходного. Вновь привести их в состояние готовности можно, лишь понизив мембранный потенциал до уровня ПП, что и является задачей К+-каналов. К+-каналы являются потенциал-зависимыми каналами с одной (активирующей) створкой. Относительно медленно открываясь (рис. 3.11, точки 2 —3), они обеспечивают выходящий ток калия и фазу гиперполяризации ПД. Закрытие створки происходит после «прохода» порогового уровня для запуска ПД (точки 4 —5). При этом скорость закрытия также невелика. В результате повышенная проницаемость мембраны для ионов К+ сохраняется еще некоторое время. Это позволяет разности потенциалов на мембране достичь уровня ПП, а часто — и опуститься ниже, что вызывает следовую гиперполяризацию. Если потенциал на мембране все же остается выше ПП, наблюдается следовая деполяризация. Оба эти состояния в дальнейшем затухают за счет деятельности постоянно открытых ионных каналов. Одновременно при проходе уровня порога ПД совершают обратное движение створки Na+-каналов. Сначала вновь быстро срабатывает А-створка, закрывая проход канала (рис. 3.11, точка 4), затем возвращается в исходное положение И-створка (створка 5). В результате предотвращается возможность входа дополнительной порции Na+ при возврате канала в состояние готовности. Пока И-створки Na+-каналов не открылись, генерация новых ПД невозможна. Это состояние называют абсолютной рефрактерностъю, по длительности оно практически совпадает с длительностью ПД. Наличие абсолютной рефрактерности не позволяет отдельным ПД накладываться друг на друга и ограничивает максимальную частоту разрядов нервных клеток величиной 500—700 Гц (реже — до 1000 Гц). Вслед за стадией абсолютной рефрактерности наступает относительная рефрактерность: лишь часть Na+-каналов вернулась в состояние готовности, еще не закрыты все потенциал-зависимые К+-каналы. В таких условиях запуск ПД возможен, но только под действием более мощного стимула. Интересным вариантом генерации ПД являются пейсмекерные нейроны (клетки-пейсмекеры). Они обладают большой постоянной проницаемостью мембраны для ионов Na+. В результате у клеток-пейсмекеров не существует стабильного ПП. Разность потенциалов на их мембране постоянно стремится вверх. Когда она достигает порогового значения, происходит запуск ПД. После ПД заряд внутри клетки оказывается на довольно низком уровне, ПП вновь растет и запускается следующий ПД, в целом же наблюдается ритмический рисунок разрядов (рис. 3.12). Рост заряда 1 внутри клетки, связанный с большим током утечки Na+, приводит к периодической самопроизвольной генерации потенциалов действия. Нейроны-пейсмекеры находятся в дыхательном центре продолговатого мозга, сходные же свойства имеют клетки центра сердечного автоматизма.
Существуют химические соединения — специфические блокаторы потенциал-зависимых Na+-каналов. Введение любого из этих препаратов блокирует вход ионов Na+ и прекращает генерацию потенциалов действия. Наиболее известный блокатор — тетродотоксин, яд рыбы-собаки (иглобрюха), обладающий очень высокой активностью. Признаками отравления являются быстрое понижение тонуса сосудов (расслабление гладких мышц в их стенках), угнетение дыхания, паралич скелетной мускулатуры. Часть молекулы тетродотоксина входит в устье потенциал-зависимого Nа+-канала и застревает в нем. В результате возникает хорошо подогнанная «пробка» и ПД не развивается. Тетродотоксин эффективен только при действии с наружной стороны мембраны (рис. 3.13). Особая группа фармакологических препаратов — местные анестетики: используется для предотвращения генерации и распространения ПД в периферических нервах. Это позволяет устранить локальные болевые ощущения. Молекулы местных анестетиков (новокаин, лидокаин) слишком велики, чтобы войти в Na+-канал снаружи; они перекрывают его, проникнув изнутри клетки либо предварительно растворившись в липидной мембране. С внутренней стороны канала действуют также блокаторы И-створки. Примером может служить батрахотоксин, яд лягушек-листолазов. Он резко увеличивает проницаемость мембран для ионов Na+, приводя к их деполяризации и нарушению сердечной проводимости.
Тетраэтиламмоний способен блокировать потенциал-зависимые К+-каналы. В его присутствии наблюдается резкое удлинение нисходящей фазы ПД. В результате возрастает и суммарная длительность потенциала действия. В настоящее время известен не один, а несколько типов потенциал-зависимых Na+- и К+-каналов. Они различаются порогами срабатывания, а также скоростью открытия и закрытия. Кроме того, в суммарные ионные токи, возникающие при развитии ПД, значительный вклад вносят ионы Ca2+. Существуют специфические потенциал-зависимые каналы, через которые Ca2+ входит в клетку; при этом он выполняет две функции: как положительно заряженный ион деполяризует мембрану и как химический агент влияет на работу других ионных каналов, ферментов, рецепторов. Важно также то, что всякий ПД уменьшает существующую разность концентраций ионов Na+ и К+ внутри и снаружи клетки. Если заставить нейрон генерировать ПД с максимальной частотой, то уже через несколько тысяч импульсов за счет выхода К+ и входа Na+ общий заряд цитоплазмы существенно приблизится к нулю. В результате И-створки Na+-каналов не смогут открыться в конце нисходящей фазы ПД и потенциалы действия прекратятся. Теперь для восстановления работоспособности необходим отдых, и тогда Na+, К+-насосы вернут разность потенциалов к уровню ПП. Для реальных режимов деятельности ЦНС подобные ситуации нетипичны: «рабочая» частота нейронов редко превышает 50—70 Гц, причем интервалы активации невелики (десятые доли секунды). Однако в случае электрошока или эпилептического припадка временное выключение импульсной активности нервных клеток весьма характерно.
|
||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 459; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.32.238 (0.008 с.) |