Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Постсинаптические потенциалы. Распространение потенциала действия по нейронуСодержание книги
Поиск на нашем сайте
Запуск импульсной активности в нервной системе осуществляют два основных фактора. Первый из них — стимулы, действующие на чувствительные клетки сенсорных систем и изменяющие проницаемость их мембраны. Это приводит к развитию особых рецепторных потенциалов и в итоге — к генерации ПД. Второй фактор — выделение медиатора из пресинаптического окончания. Попав в синаптическую щель, медиатор воздействует на постсинаптическую мембрану, возбуждая или тормозя следующий нейрон. Процессы подобного возбуждения или торможения связаны с деятельностью еще одного типа ионных каналов — лиганд-зависимых (хемочувствительных). Они находятся на мембране, непосредственно окружающей синаптический контакт. Обычно они закрыты. Их открывание происходит лишь при появлении медиатора, несущего сигнал химического вещества (отсюда термин «хемочувствительные»). Лиганд-зависимые каналы можно разделить на три основных класса: избирательно проницаемые по отношению к ионам Na+, ионам К+ и ионам Cl−. Отрывание первых из них приведет к входу в клетку ионов Na+ и деполяризации нейрона (рис. 3.14, а), во время которой разность потенциалов на мембране оказывается приближенной к порогу запуска ПД. В этот момент меньший, чем обычно, стимул может вызвать реакцию нейрона, т. е. нервная клетка находится в относительно возбужденном состоянии. В связи с этим локальная деполяризация мембраны под действием медиатора была названа возбуждающим постсинаптическим потенциалом (ВПСП). Медиаторы, вызывающие ВПСП, отнесены к группе возбуждающих медиаторов.
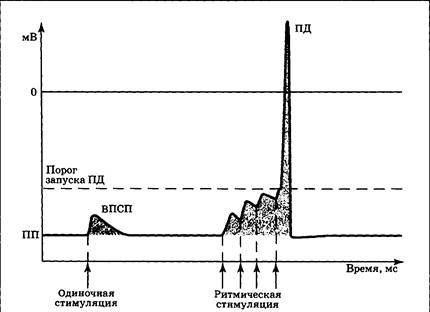
Открывание хемочувствительных Cl−-каналов приводит к входу в клетку ионов хлора; открывание К+-каналов — к выходу ионов калия. В этих случаях возникает гиперполяризация и разность потенциалов на мембране нейрона увеличивается по абсолютной величине (рис. 3.14, б), поэтому для запуска ПД необходим больший, чем обычно, стимул. Следовательно, нервная клетка находится в относительно заторможенном состоянии. В связи с этим локальная гиперполяризация мембраны под действием медиатора была названа тормозным постсинаптическим потенциалом (ТПСП). Медиаторы, вызывающие ТПСП, отнесены к группе тормозных медиаторов. Усредненные параметры ВПСП и ТПСП весьма близки (рис. 3.14). Их длительность составляет обычно около 10 мс (иногда 50—100 мс), что существенно больше, чем в случае ПД. Амплитуда ВПСП и ТПСП определяется длительностью и крутизной наклона их первой фазы, которая зависит от количества и длительности существования медиатора в синаптической щели. Амплитуда одиночных постсинаптических потенциалов в ЦНС составляет 1—5 мВ. В крупном нервно-мышечном синапсе аналог ВПСП — потенциал концевой пластинки, может достигать 40 и более мВ. При детальном анализе сигнала можно видеть, что первые фазы ВПСП и ТПСП имеют ступенчатый характер, т. е. нарастают дискретно, шагами (квантами). Такая дискретность связана с тем, что выброс медиатора в синаптическую щель так-же идет квантами, где квант — одна везикула. В каждой везикуле содержится несколько тысяч молекул медиатора, и их воздействие на постсинаптическую мембрану вызывает сдвиг потенциала примерно на 0,1 мВ. В подавляющем большинстве случаев (кроме потенциала концевой пластинки) одиночный ВПСП не способен запустить ПД, так как возбуждение, вызываемое медиатором, не дорастает до порогового уровня. Для достижения порога запуска ПД необходима суммация (наложение) нескольких ВПСП. Выделяют два варианта суммации — временную и пространственную. Временная суммация — объединение эффектов стимулов, пришедших по одному «каналу» с большой частотой (рис. 3.15): если к еще не угасшему ВПСП присоединить второй, затем третий и т.д., — возникнет реальная возможность запустить ПД. Это означает, что сигнал, достигший синапса, достаточно интенсивен и «заслуживает» того, чтобы быть переданным дальше по сети нейронов.
Пространственная суммация заключается в наложении друг на друга ВПСП соседних синапсов 1 — 3 в некоторой близлежащей точке постсинаптической мембраны 4 (рис. 3.16), обладающей потенциал-зависимыми ионными каналами. Схема пространственной суммации напоминает логическую ячейку по типу «И», т. е. результат положительный, если несколько условий будут выполнены (несколько входных сигналов одновременно достигнут нервной клетки). В ходе деятельности нейронов эффекты пространственной и временной суммации объединяются, и чем больше синапсов участвуют в этом процессе (срабатывают относительно одномоментно), тем больше вероятность достичь порога запуска ПД. При этом часть синапсов может обладать тормозными свойствами и вызывать ТПСП, вычитающиеся из суммы возбуждающих влияний. В результате в первом приближении условие запуска ПД в каждый момент времени можно определить следующим образом: ПП + (сумма всех ВПСП) − (сумма всех ТПСП) > порог запуска ПД Оценить вклад конкретных постсинаптических потенциалов в этот процесс непросто. Дело в том, что их влияние быстро затухает по мере удаления от места возникновения. Кроме того, затухание в отростках происходит быстрее, чем в теле нейрона, а также тем быстрее, чем тоньше отросток. И наконец, мембрана нейрона в разных местах имеет разную возбудимость. Она максимальна в аксональном холмике (место отхождения аксона от тела) и в местах первого ветвления крупных дендритов. В итоге оказывается, что чем ближе конкретный синапс к этим точкам, тем больше его вклад в управление генерацией ПД. Одного ТПСП, возникшего рядом с аксональным холмиком, может оказаться достаточно для прекращения проведения сигнала (логическая ячейка по типу «НЕ»). Процесс суммирования ВПСП и ТПСП, возникших в разных синапсах, является основной вычислительной операцией, протекающей на нейронах ЦНС. При ее реализации сигналы имеют возможность «подтвердить» свою значимость, могут объединиться с другими сигналами и сформировать некоторый «информационный образ», могут быть заблокированы (при наличии определенных условий — сигналов по тормозным каналам). В связи с этим способность ЦНС выполнять сложные вычислительные операции определяется не ее общим весом и даже не числом нейронов, а именно количеством синапсов. По мере развития мозг способен формировать дополнительные синапсы, увеличивая свои потенциальные возможности. Особенно интенсивно этот процесс идет в раннем детстве, когда нервная система настраивается на предстоящий уровень информационной нагрузки. Если ПД возник в некоторой точке 1 мембраны (рис. 3.17, а), он начинает выполнять функцию запускающего стимула по отношению к соседним областям. При этом ток натрия, формирующий первую фазу ПД, оказывает действие, аналогичное влиянию деполяризующего электрического стимула. В результате потенциал-зависимые ионные каналы на еще не активированной мембране начинают открываться, и ПД делает первый шаг. Развившись в точке 2, ПД запускает возбуждение в точке 3 и т. д. Этот процесс можно уподобить волне, расходящейся по воде от места падения камня. В ходе этого распространения фронт ПД движется от точки возникновения в разные стороны без «возвратов» назад, так как только что сработавшая мембрана приходит в состояние абсолютной рефрактерности. В каждый момент времени она может проводить сигнал только в одном направлении, и повторное проведение возможно лишь после возврата потенциал-зависимых каналов в состояние готовности.
Необходимость открывания ионных каналов на последовательных участках мембраны делает проведение ПД весьма медленным — около 0,5—1 м/с (рис. 3.17, б), при этом ширина одномоментно активированной зоны (фронт ПД) составляет примерно 1 мм. Скорость распространения можно увеличить, делая нервные волокна более толстыми: в гигантском аксоне кальмара при толщине 1 мм она достигает 10 м/с. Другой путь — формирование миелиновых оболочек. В этом случае очередным возбужденным участком при проведении ПД будет не соседняя точка мембраны, а ближайший перехват Ранвье. В результате импульс будет распространяться прыжками (сальтаторно; рис. 3.17, в), развиваясь только на участках аксона, не закрытых Шванновскими клетками. Ширина таких участков — 0,5 мкм (против 1 мм для миелинового сегмента). Скорость «прыжка» определяется толщиной миелиновой оболочки. При общем диаметре волокна 1 мкм она составляет 3 м/с, при максимальном диаметре 22 мкм — 120 м/с. Кроме резкого увеличения скорости проведения, сальтаторный вариант является очень экономичным, поскольку изменения ионного баланса в результате ПД происходят только в зоне перехватов. Скорость, близкая к 100 м/с, характерна для передачи кожной и мышечной чувствительности, двигательных команд. Именно задержки, возникающие при такой передаче, определяют латентный (скрытый) период простых сенсомоторных реакций. Он составляет около 20 мс, что соответствует «длине пути» нервного импульса — около 2 м (по руке к спинному мозгу и обратно). Конечной целью бегущего по нерву ПД являются пресинаптические окончания, где электрический сигнал превращается в химический, и скачок потенциала на мембране запускает выделение медиатора, т. е. пока сигнал остается в «границах» одного нейрона, он передается в электрической форме (ПД). Для передачи между нейронами информация преобразуется в химическую форму (медиатор). Подействовав на постсинаптическую мембрану, медиатор вновь переводит сигнал в электрическую форму (ВПСП, ТПСП, запуск ПД). Все это свидетельствует о тесном взаимодействии электрических и химических процессов в ходе реализации различных функций мозга.
|
||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 367; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 52.14.49.59 (0.009 с.) |