Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Безусловное и условное торможениеСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
В нервной системе можно наблюдать два базовых процесса — возбуждение и торможение. На уровне нервной клетки возбуждению соответствует уменьшение мембранной разности потенциалов (деполяризация), а торможению — ее увеличение (гиперполяризация). Торможение, как и возбуждение, — активный (т. е. требующий затраты энергии) процесс, связанный с выбросом тормозных медиаторов, таких, как ГАМК, глицин. На уровне поведения торможение проявляется в виде ослабления либо полного прекращения двигательных и вегетативных реакций. Развивая идеи И. М. Сеченова, И. П. Павлов выделил два типа торможения — безусловное и условное. К первому из них, по аналогии с безусловными рефлексами, он отнес не нуждающиеся в выработке врожденные видоспецифичные формы торможения. Различные варианты условного торможения, как и условные рефлексы, вырабатываются в течение жизни, индивидуальны и требуют специального обучения. Безусловное торможение Павлов подразделил на внешнее и запредельное (табл. 4.3). Внешнее торможение впервые было описано в ситуациях, когда неожиданные посторонние сигналы угнетали выработанные в ходе экспериментов условные рефлексы. На такие раздражители (громкий звук, сильный запах) у собак возникал ориентировочный рефлекс, на фоне которого остальная деятельность временно прекращалась. У человека подобное явление называется отвлечением. Именно из-за внешнего торможения отсутствие помех является одним из условий успешной выработки условных рефлексов. Биологический смысл внешнего торможения — остановка текущей деятельности организма и реализация исследовательского поведения, поскольку из внешней среды может поступить жизненно важная информация. Ситуация внешнего торможения иллюстрирует проявление одного из важнейших принципов работы мозга — принципа доминанты. Учение о доминанте было разработано А. А. Ухтомским (1875—1942), обратившего внимание на то, что, несмотря на все многообразие возможных реакций, в ЦНС в каждый момент времени может быть выделен один главный, господствующий очаг возбуждения («ведущий» безусловный рефлекс). Этот очаг и связанная с ним деятельность были названы доминантой. Доминантная область мозга через тормозные интернейроны может снижать возбудимость других аналогичных центров. Более того, направлявшиеся в эти центры сигналы начинают с большей вероятностью возбуждать доминантный очаг, т. е. поступающая в ЦНС информация будет анализироваться с точки зрения соответствующего типа деятельности. Например, если мы очень голодны, все сенсорные сигналы рассматриваются как несущие информацию о возможном удовлетворении пищевой потребности.
В каждый момент времени существует только одна явная доминанта, и в ЦНС происходит постоянная конкуренция различных безусловных рефлексов, в результате которой побеждает наиболее возбужденный центр. Победа эта только временная, и в случае необходимости нервная система способна оперативно менять (переключать) доминанту. Торможение условно-рефлекторного слюноотделения собаки при громком звуке, прекращение плача ребенка при виде яркой игрушки — все это примеры смены доминанты. Запредельное торможение возникает в нервной системе в ответ на сильные или длительно действующие раздражители, когда наступает предел работоспособности (утомление) нервных клеток. Этот вид торможения Павлов назвал также охранительным (защитным). Перевозбуждение человека, постоянный шум и яркий свет, длительная и интенсивная умственная деятельность могут привести к состоянию заторможенности. Конкретные физиологические причины запредельного торможения могут быть разными: включение особых тормозных механизмов, ограничивающих возбуждение в нейросетях, недостаток питания (глюкозы) и кислорода, накопление отходов обмена веществ, истощение запасов медиаторов в синапсах, нарушение ионного баланса в нервной ткани. При этом утомление тормозных нейронов часто наступает раньше, чем активационных. В результате первая стадия запредельного торможения может иметь парадоксальный характер: наблюдается всплеск плохо контролируемой активности, эмоций (капризы и плач маленького ребенка перед засыпанием), после чего происходит общее торможение деятельности мозга. Самый естественный способ устранения запредельного торможения — отдых и сон. Если запредельное торможение велико, сон наступает очень быстро. Клиническим вариантом запредельного торможения является летаргический сон, возникающий в результате тяжелых психических травм, причем больной может находиться в таком состоянии в течение недель и месяцев. Перейдем к условному торможению, т. е. к ситуации, когда у животного (человека) вначале выработан условный рефлекс, а затем происходит его модификация. Павлова гораздо больше интересовал именно этот вид торможения, поскольку он вызывает изменение существующих временных связей. Выделено четыре основных варианта условного торможения — угасательное, дифференцировочное, условный тормоз и запаздывательное. Угасательное торможение (угашение условного рефлекса) развивается в том случае, когда при уже выработанной временной связи условный раздражитель перестает сопровождаться подкреплением. В итоге рефлекторный ответ (рис. 4.22) постепенно уменьшается по S-образной кривой, очень сходной с кривой обучения. Чем чаще предъявляется неподкрепляемый условный раздражитель, тем быстрее идет угашение. Чем прочнее условный рефлекс, тем хуже он угашается (оборонительный медленнее, чем пищевой). Важно, что при угасательном торможении происходит не разрушение временной связи, а лишь ее блокирование, переход в скрытое состояние. Действительно, если на фоне уже произведенного угашения подать подкрепление, то реакция проявится в полном объеме уже при следующем включении условного стимула. Павлов назвал явления такого рода растормаживанием.
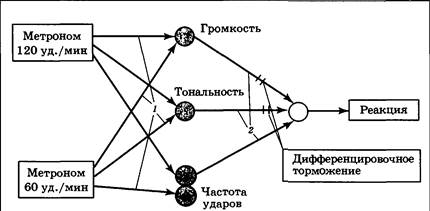
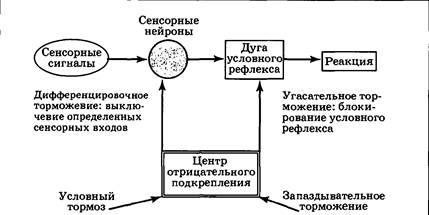
Нейрофизиологическая основа угашения (и условного торможения вообще) связана с деятельностью центра отрицательного подкрепления. Активация этого центра происходит при получении отрицательного подкрепления, а также при неполучении ожидаемого положительного подкрепления. В последнем случае к дуге уже сформированного условного рефлекса присоединяется «тормозной блок», представляющий собой корковый тормозный интернейрон, «обучающийся» реагировать на активацию центра отрицательного подкрепления. При растормаживании центр положительного подкрепления по принципу доминанты выключает центр отрицательного подкрепления, что приводит к снятию торможения с временной связи. Угашение — биологически важное приспособление, благодаря которому организм перестает напрасно тратить энергию и реагировать на сигнал, потерявший свое значение. В ходе угасательного торможения идет отрицательное обучение, переводящее временные связи (память) в скрытое состояние с тем или иным уровнем доступности. При длительном неиспользовании условных рефлексов идет их самопроизвольное угашение («забывание»). Второй вид условного торможения — дифференцировочное. Оно вырабатывается при противопоставлении двух сходных сенсорных агентов — постоянно подкрепляемого и неподкрепляемого. В эксперименте Павлова вначале был образован пищевой слюноотделительный условный рефлекс на стук метронома, настроенного на частоту 120 ударов/мин. Если затем предъявить метроном с другой частотой, например 60 ударов/мин, он также будет вызывать условную реакцию. В дальнейшем производилось чередование подкрепляемого метронома 120 ударов/мин и метронома 60 ударов/мин, чье включение не подкреплялось. После 20—30 таких «парных» стимуляций оказывается, что собака начинает различать (дифференцировать) один метроном от другого; реакция на первый из них сохраняется, на второй — исчезает. В отличие от ситуации угасательного торможения, в данном случае рефлекс на исходный условный стимул сохраняется. Суть происходящего заключается в сенсорной перенастройке мозга. Центр, воспринимающий условный сигнал, начинает работать более тонко, избирательно и различать информационные каналы, по которым поступает возбуждение после применения первого и второго метрономов. Задача эта непростая, так как степень сходства двух используемых стимулов очень высока: применяется одинаковый по громкости, высоте и другим характеристикам звуковой щелчок (звук метронома), и разница заключается лишь в частоте следования щелчков, поэтому подключающийся в данном случае к рефлекторной дуге «тормозный блок» должен быть установлен в сенсорных центрах и локально блокировать те сенсорные входы, которые являются общими для обоих метрономов (рис. 4.23). Чем ближе сенсорные характеристики дифференцируемых сигналов, тем длительнее и тяжелее для нервной системы процедура выработки различения. При некотором уровне сходства дифференцировка вообще невозможна: различение звуковых тонов, отличающихся на 1/8 нотного ряда, возможно для собак, но далеко не для всех людей. У крыс бессмысленно вырабатывать дифференцировку на частоту ударов метронома, но легко выработать различение звука по громкости.
Дальнейший анализ явления дифференцировочного торможения показал, что оно имеет более широкий спектр проявлений, чем предполагалось изначально, и участвует в образовании любого условного рефлекса. Действительно, на начальной стадии обучения формирование временных связей происходит между корковым представительством безусловного рефлекса и множеством разномодальных сенсорных центров коры, т. е. формируется большое количество обстановочных условных рефлексов. Павлов назвал это стадией генерализации условного рефлекса. Так, в случае условной реакции активного избавления (см. разд. 4.4) на начальной стадии обучения само помещение в экспериментальную камеру вызывает у крысы прыжок на полку, иными словами, действия зрительных, обонятельных, тактильных обстановочных стимулов оказывается вполне достаточно для запуска реакции. Приходится специально угашать эти рефлексы, раз за разом снимая крысу с полки и опуская ее на пол камеры. При этом условный стимул (звонок) продолжает подкрепляться. В результате дифференцировочное торможение «убирает» временные связи, сформировавшиеся между центрами неслуховых сенсорных систем и двигательным центром. При дальнейшем обучении дифференцировка затронет уже слуховую систему, и постепенно можно добиться, чтобы прыжок не запускался иными звуками, кроме звонка. В результате из множества изначально сформировавшихся реакций сохранится только одна, т. е. мозг окажется специально настроенным реагировать на условный стимул (по Павлову — наступит стадия специализации условного рефлекса). Как генерализация, так и специализация условных рефлексов имеют достаточно очевидный биологический смысл. Попав в новую ситуацию и получив подкрепление в первый раз, особь не знает, какие из воздействовавших на нее сенсорных стимулов действительно значимы (сигнализируют о появлении подкрепления), а какие — случайны. В таком положении формирование временных связей идет по избыточному принципу: устанавливаются ассоциации между всеми активированными центрами. Хотя качество деятельности еще невелико, происходит быстрая модификация поведения в зависимости от условий среды. Все это особенно важно, когда особи угрожает опасность (очевидно, что лучше выполнить лишнюю оборонительную реакцию, чем погибнуть из-за того, что эта реакция еще не сформирована). В дальнейшем по мере повторов ситуации случайные раздражители появляются редко, а значимые — стабильно; кроме того, случайные раздражители могут встретиться и в других ситуациях, причем подкрепления вслед за ними не последует. Все это будет вести к укреплению одних временных связей, угашению и дифференцировочному торможению других, иначе — к переходу к более «экономичному» специализированному варианту организации условно-рефлекторного ответа. Третий тип условного торможения называется условный тормоз. В экспериментах Павлова он вырабатывался следующим образом: к условному раздражителю (например, включение лампочки) присоединялся еще один исходно индифферентный сигнал (звонок), и это сочетание не подкреплялось; в то же время одиночный условный раздражитель продолжал подкрепляться. Иными словами, проводилось чередование подкрепляемой лампочки и неподкрепляемого комплекса звонок плюс лампочка. Оказалось, что исходно собака реагирует на этот комплекс, но по мере проведения обучения реакция ослабевает и прекращается. Следовательно, звонок (он и был определен как «условный тормоз») становится специфическим сигналом, чье появление предотвращает реакцию на условный раздражитель. Характерно также то, что хорошо отработанный условный тормоз при первом же использовании перед каким-либо другим условным рефлексом ослабляет или даже полностью блокирует его. Следовательно, в отличие от двух предыдущих случаев, когда влияние отрицательного обучения распространялось только на «свою» временную связь, условный тормоз способен вызывать достаточно генерализованное торможение всей сети приобретенных рефлексов. Это позволяет предположить, что в данном случае процедура обучения (увеличения эффективности синапсов) происходит на входе в центр отрицательного подкрепления, который, в свою очередь, при появлении условного тормоза блокирует широкий круг функций головного мозга (рис. 4.24).
Данный тип условного торможения встречается довольно часто и в обычной жизни: это ситуации, связанные с запретом в определенных условиях какой-либо деятельности. Например, кошка в присутствии хозяина не залезет на стол с едой (вид еды — условный раздражитель; наличие хозяина — условный тормоз пищевой реакции). Роль мощного условного тормоза играет у человека (ребенка) слово «нельзя». При дрессировке служебных собак стремятся выработать остановку всякого текущего поведения на команду «фу». Четвертый вид условного торможения — запаздывательное. Оно проявляется при выработке отставленных условных рефлексов, т. е. в случаях, когда между условным стимулом и подкреплением создается значительный интервал. В экспериментах Павлова исходно этот интервал составлял 10—20 с, пища давалась не на фоне действия условного стимула, а после его выключения. Выработав условный рефлекс, интервал начинали увеличивать, доводя до двух минут. Если такое увеличение осуществлять достаточно медленно, рефлекс не разрушается, и можно наблюдать, что в первые 1—1,5 мин после условного стимула у собак развивается заторможенное состояние. Некоторые животные даже на короткое время засыпали, просыпаясь ближе к моменту получения подкрепления и демонстрируя условно-рефлекторное слюноотделение. Таким образом, данный тип условного торможения характерен для ситуаций ожидания, когда мозг должен некоторое время находиться в состоянии готовности к деятельности, затрачивая при этом минимум энергии. В природе проявления запаздывательного торможения чрезвычайно свойственны хищникам, охотящимся из засады (кошачьи). В этом случае, как и в случае условных рефлексов на время, мы вновь встречаемся со способностью нервной системы «отсчитывать» временные интервалы — на этот раз между условным стимулом и подкреплением; именно на этот интервал происходит блокирование временной связи. Важно также то, что запаздывательное торможение снижает уровень активности всей нервной системы, действуя, как и в случае условного тормоза, на входе в центр отрицательного подкрепления (рис. 4.24). Переход в состояние общей заторможенности и сна очень характерен для людей, длительное время находящихся в положении ожидания какого-либо события (сигнала). Итак, мы рассмотрели четыре вида условного торможения, суть которых состоит в модификации уже имеющихся условных рефлексов. Система подобного отрицательного обучения — необходимое дополнение системы положительного обучения, т. е. собственно процедуры выработки условных рефлексов; оба этих процесса необходимы для гибкой, адекватной адаптации поведения к меняющимся условиям внешней среды. Условное торможение является в большинстве случаев даже более сложной задачей и в ходе эволюции развивается позже, чем способность к образованию ассоциативных связей. Система сна и бодрствования Возбуждение и торможение, возникнув в определенном участке нервной системы, способны распространяться. Этот процесс, открытый еще в XIX веке, получил название иррадиации. Так, если раздражать электрическим током двигательную область коры, то постепенно в ответ вовлекается все большее количество мышц, а затем реакция выходит за пределы двигательной системы, например, повышается слюноотделение и учащается сердцебиение. Зная механизмы работы мозга на клеточном уровне, легко понять, что в основе иррадиации лежит распространение ПД по аксонам возбуждающих либо тормозных нейронов. Учтем также процессы суммации, когда, повышая возбудимость синапсов, повторные нервные сигналы «прокладывают» себе путь, все более удаляясь от точки возникновения. При снижении уровня возбуждения (торможения) наблюдается противоположный по отношению к иррадиации феномен — концентрация (сосредоточение) нервных процессов в исходном пункте. Важно, что при концентрации в определенном участке мозга нервный процесс одновременно вызывает (индуцирует) противоположный нервный процесс в окружающих зонах. В том случае, когда возбуждение индуцирует торможение, говорят об отрицательной индукции. В обратном случае, когда торможение индуцирует возбуждение, индукция положительная. Особенно важное биологическое значение имеет отрицательная индукция. Смысл ее таков: сильный очаг возбуждения способен тормозить другие аналогичные очаги в нервной системе (так реализуется принцип доминанты). Наиболее широкие процессы иррадиации возбуждения и торможения в ЦНС обеспечивают системы сна и бодрствования. У человека выявлено большое число физиологических показателей, которые изменяются с периодом в 24 ч (температура тела, артериальное давление, выделение гормонов), причем такая ритмичность сохраняется даже при неизменности всех внешних факторов (например, если человек долгое время находится в пещере). Это говорит о том, что в процессе эволюции в условиях планеты Земля, совершающей оборот вокруг своей оси за 24 ч, организмы приспособились к такому ритму и появились эндогенные (внутренние) механизмы, обусловливающие его сопровождение. Запускаемые ими ритмы принято называть околосуточными, или циркадианными (лат. circa — около и dies — день). Наиболее выражен суточный цикл сон — бодрствование. Он реализуется как за счет безусловно-рефлекторной основы, главные компоненты которой относятся к группе витальных врожденных реакций, так и за счет определенных адаптивных процессов, которые и будут рассмотрены ниже. Состояние бодрствования характеризуется высокой нервной активностью. В это время происходит взаимодействие организма с внешней средой, формирование и реализация различных рефлекторных реакций. Бодрствующее состояние поддерживается особыми центрами бодрствования. Сон — это физиологическое состояние, при котором значительно снижается реакция нервной системы и всего организма на внешние раздражители. Состояние сна является следствием включения особых центров сна головного мозга и сопровождается снижением интенсивности физиологических процессов, общей неподвижностью. Павлов предлагал выделять два вида сна — активный и пассивный. Первый он определял как возникающий при отсутствии (недостатке) внешних раздражителей. Соответственно для поддержания бодрствующего состояния часто необходим некоторый минимум сенсорных стимулов. Активный сон, по Павлову, возникает как результат работы особых центров сна. Павлов представлял активный сон в виде тормозного процесса, иррадиирующего из определенного утомленного центра коры и захватывающего весь головной мозг. Дальнейшие исследования показали, что гораздо чаще источником сна являются другие, более глубоко расположенные структуры головного мозга. Кратко охарактеризуем основные центры бодрствования и сна головного мозга (рис. 4.25).
Функции главного центра бодрствования нашего мозга выполняют в ЦНС ретикулярные ядра моста. Каждая сенсорная система, помимо передачи информации в специфические зрительные, слуховые, вкусовые зоны, часть сигналов направляет именно сюда. В ретикулярных ядрах моста (ретикулярная формация моста) происходит их суммирование и оценка общего уровня сенсорного «давления» на организм. Далее сигналы от ретикулярных ядер чрезвычайно широко иррадиируют по ЦНС. Анатомическую основу для этого обеспечивают коллатерали аксонов их нейронов, которые образуют возбуждающие синапсы на самых разных уровнях — от коры больших полушарий до серого вещества спинного мозга. Таким образом, чтобы вывести мозг из сонного состояния, достаточно подать сильный сенсорный сигнал (звонок будильника), а для ускорения утреннего пробуждения и вхождения в «рабочий режим» полезны яркий свет или сильные вкусовые ощущения. Чем больше информации поступает из внешней среды, тем активнее функционирует и откликается на нее мозг. Однако при переизбытке сигналов работоспособность нервной системы может истощиться, и тогда возникнет охранительное запредельное торможение. С другой стороны, при недостатке раздражителей уровень бодрствования упадет и станет возможным переход в состояние пассивного сна. В качестве ключевых центров сна рассматриваются содержащие серотонин структуры среднего мозга и моста (ядра шва) и влияющее на них центральное серое вещество среднего мозга. Функции серотонина как медиатора нервной системы уже обсуждались в разделе, посвященном общей физиологии нервной системы. Напомним, что они связаны с тормозной модуляцией сенсорных входов. Как и в случае ретикулярных ядер моста, коллатерали аксонов серотонинергических нейронов обнаруживаются на всех уровнях ЦНС. Между ретикулярными ядрами моста и центрами сна существуют антагонистические отношения, являющиеся частным случаем проявления принципа доминанты. При этом глутаматергические нейроны центров бодрствования нуждаются в посреднике для реализации тормозного действия на центры сна. Роль такого посредника выполняет голубое пятно — область норадренергических нейронов в верхней части моста. Стимуляция этой зоны вызывает выключение центров сна и пробуждение. Следующей важной областью, связанной с регуляцией суточных ритмов, являются супрахиазменные ядра гипоталамуса. Контактируя со зрительной системой, они оказываются настроенными на суточные колебания освещенности в окружающей среде. В результате часть нервных клеток этой области обнаруживает активность в темноте, а часть — в светлое время суток. Первые посылают свои аксоны в центры сна, вторые — в центры бодрствования. Нейроны супрахиазменных ядер обладают свойством автоматизма (будучи изолированными от всего остального мозга, продолжают поддерживать суточный ритм). Их назначение состоит в том, чтобы вовремя сообщить ретикулярным ядрам моста, что уже «пора вставать», а центральному серому веществу и ядрам шва — что уже «пора спать». Здесь, как и в случае рефлексов на время и запаздывательного условного торможения, мы сталкиваемся с присущей нервным структурам способностью оценивать и отсчитывать временные интервалы. Какие процессы лежат в ее основе? Если речь идет о коротких временных интервалах (секунды — десятки секунд), мы, как правило, имеем дело с нейронами-пейсмекерами либо замкнутыми нейронными контурами, по которым с определенным периодом циркулирует возбуждение. Если это десятки минут и часы (как в случае супрахиазменных ядер), то можно говорить о некоторых внутриклеточных ритмах, когда в роли счетчика времени выступают цепочки биохимических реакций. Несмотря на свойство автоматизма, супрахиазменные ядра способны перенастраиваться под действием навязанного извне суточного ритма активности (освещенности). Это оказывается особенно необходимым при резкой смене часовых поясов, когда день превращается в ночь, а ночь — в день. Перенастройка занимает до двух недель. Частая смена часовых поясов, отсутствие стабильного режима дня, круглосуточное бодрствование ухудшает состояние супрахиазменных ядер, что может привести к серьезным сбоям в ее функционировании, проблемам с засыпанием, бессоннице. Последняя из представленных на рис. 4.25 структур — ретикулярные ядра продолговатого мозга. Не являясь самостоятельными центрами сна, они посылают в центральное серое вещество сигналы, связанные с оценкой содержания в крови различных важных для деятельности мозга веществ (глюкозы, токсинов, отходов обмена веществ). За счет активации ретикулярной формации продолговатого мозга сонное состояние может развиться после еды, при болезнях, физическом утомлении. В целом можно заключить, что центры сна и бодрствования находятся в постоянных конкурентных отношениях и степень доминирования тех или других зависит от многих факторов: времени суток, сенсорной нагрузки, физического состояния. Это позволяет нервной системе гибко варьировать общий уровень своей активности в зависимости от значимости текущей деятельности (ожидаемого подкрепления) и степени утомления. В результате ЦНС, с одной стороны, функционирует в режиме максимальной экономии ресурсов, с другой — в случае необходимости может быстро переходить в состояние высокой работоспособности, быстрого и адекватного реагирования на внешние стимулы. С помощью электрофизиологических методик установлено, что естественный ночной сон состоит из нескольких повторяющихся циклов. В течение каждого такого цикла наблюдаются закономерные изменения в электроэнцефалограмме (ЭЭГ) — электрической активности коры больших полушарий. Две основные характеристики ЭЭГ — это частота и амплитуда составляющих ее волн. Известно, что в зависимости от функционального состояния человека характер ЭЭГ меняется: чем больше активность мозга, тем больше частота и ниже амплитуда волн ЭЭГ. В спокойном бодрствующем состоянии в ЭЭГ преобладает альфа-ритм (8—12 Гц), при активной умственной деятельности он сменяется бета-ритмом (13—30 Гц). Во время засыпания активность коры снижается и в ЭЭГ появляются сначала тета-волны (4—7 Гц), а затем дельта-волны (1—3 Гц) (рис. 4.26). Сон с преобладанием медленных дельта-волн является основной фазой сна (около 80% общей его длительности) и называется медленным сном. В ходе медленного сна наблюдается глубокое торможение большинства областей ЦНС, происходит «физиологический отдых» — восстановление запасов химических веществ и энергии, затраченных во время бодрствования.
Можно наблюдать, что в течение ночи медленный сон 4—5 раз периодически сменяется другой фазой сна. В это время дельта-волны замещаются на быструю низкоамплитудную активность, сходную с ЭЭГ в состоянии бодрствования. Кроме того, наблюдается повышение температуры, частоты дыхания и сердцебиений, артериального давления, усиление потоотделения, но порог пробуждения в этот момент оказывается еще выше, чем при медленном сне, т. е. торможение сенсорных входов достигает максимума. Происходит также исчезновение мышечного тонуса (полное расслабление мускулатуры). Совокупность данных признаков позволила назвать данную фазу сна парадоксальной или быстрым сном. Во время этой стадии можно наблюдать также быстрые движения глазных яблок — отсюда третье название: стадия быстрых движений глаз, или стадия БДГ (англ. rapid eye movement — REM-сон). Во время первого цикла (после первого интервала медленного сна) стадия БДГ еще очень короткая — примерно 5 мин. Но с каждым новым циклом ее длительность нарастает и достигает 20—25 мин (суммарно около 20% от всей длительности сна). Показано, что если человеку не позволять в течение ночи находиться в этой стадии, то утром он оказывается невыспавшимся, отмечается повышенная утомляемость и раздражительность. При длительном лишении быстрого сна могут развиться неврозы. Предполагается, что во время быстрого сна мозг осуществляет обработку информации, накопленной за прошедший период бодрствования. Судя по имеющимся данным, именно эта обработка является главной причиной сновидений. Еще Сеченов определял сновидения как «небывалые комбинации бывалых впечатлений». Действительно, если проанализировать любой сон, то можно увидеть, что он складывается из уже известных элементов. Однако в итоге сновидения могут носить не только образно-эмоциональный, но также прогностический и творческий характер, отражая все многообразие деятельности нашего мозга. Чрезвычайно характерно, что стадия БДГ у взрослых заметно сокращается по сравнению с детьми. Не исключено, что это связано с постепенным снижением новизны получаемых от жизни впечатлений. Анатомически с запуском БДГ-сна связана гигантоклеточная область ретикулярных ядер продолговатого мозга, а также часть нейронов голубого пятна. Представленные на рис. 4.25 центры обеспечивают врожденные механизмы сна и бодрствования, но, исходя из общих принципов организации ВНД, над этой системой может быть надстроено некоторое количество условных рефлексов. Так, если сон начинается в одно и то же время в постоянном месте, факторы времени и обстановки могут стать условными стимулами. Сон, возникающий при этом, как и всякая приобретенная реакция, имеет корковое происхождение и называется условно-рефлекторным. Как уже говорилось, снотворные препараты в большинстве своем являются агонистами ГАМК. Основное их действие направлено на блокаду таламо-кортикальной передачи сенсорной информации за счет усиления внутриталамического торможения. Введение агонистов ГАМК не только прерывает сенсорный приток в кору (выключение сознания), но и резко ограничивает тонус ретикулярных ядер моста, что снижает общий уровень активности в мозге и приводит к переходу к естественному сну. В норме снотворный препарат нужен только для запуска сонного состояния, однако при перевозбуждении или нарушениях функций центров сна переход к естественному сну может не произойти. В этом случае через 2—3 ч наступит пробуждение и человек будет вынужден принять следующую дозу препарата. Понятно, что в такой ситуации полноценный отдых ЦНС невозможен. Отдельным видом сна коркового происхождения является гипнотический сон. В этом случае производится длительное монотонное сенсорное воздействие. Его цель — вызвать запредельное торможение локальной корковой зоны, которое затем распространится на всю кору больших полушарий, приводя к засыпанию и выключению сознания. На этой стадии крайне важно желание пациента быть загипнотизированным, поскольку любое сопротивление с его стороны нарушит иррадиацию торможения. После засыпания гипнотизер, используя различные словесные команды, может растормозить отдельные корковые зоны и, не пробуждая пациента, вызвать у него разнообразные двигательные и речевые реакции, в том числе обычно подавляемые сознанием. Ситуация, когда большинство зон коры (в том числе центры сознания) заторможены, а часть из них — нет, наблюдается и при сомнамбулизме (снохождении). Сомнамбулизм рассматривается как вариант патологического сна, обусловленный нарушением баланса возбуждения и торможения в локальных зонах коры больших полушарий. Другой вариант патологического сна — летаргический сон (упоминался выше в связи с развитием чрезвычайно сильного запредельного торможения).
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 3394; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.117.254.177 (0.014 с.) |