

Мы поможем в написании ваших работ!
ЗНАЕТЕ ЛИ ВЫ?
|
Ареал и распространение видов в его пределах
- Принцип экологической индивидуальности видов Л. Г. Раменского — Г. А. Глизона
- Правило колебания границ ареала
- Принцип воздействия факторов В. Тишлера
- Правило соответствия вида и ценоза
- Правило географической изменчивости кружева ареала
- Правило ограничивающих факторов Правило стимулирующего действия температур В. Шелфорда — Т. Парка
- Правило лимитирующего значения крайних летних и зимних температур А. Н. Голикова — О. А. Скарлато
- Правило викариата Д. Джордана
- Правило представительства рода одним видом А. Монара
- Принцип конкурентного исключения Г. Ф. Гаузе
- Принцип видо-родового представительства И. Иллиеса
- Принцип сосуществования, или парадокс Дж. Хатчинсона
- Правило географического оптимума
| Существуют два основных способа изображения ареалов на картах: путем вычерчивания их границ в виде сплошной или пунктирной (в случае сомнений в верности проведения) линии и методом нанесения точек в географические места обнаружения особей вида. В первом случае обычно производится довольно смелая экстраполяция, как правило, путем соединения крайних точек обнаружения вида, т. е. оба способа отображения границ ареала фактически совпадают.
Если в какой-то географической точке никто никогда не вел фаунистические или флористические исследования, точек находок, естественно, не возникает. Кроме того, при многолетнем изучении широких регионов, как это приходилось делать автору в Средней Сибири, нетрудно убедиться, что некоторые птицы, например, коростель, порой фактически исчезают на краю своего ареала на ряд лет, а затем вновь появляются. Для упомянутого вида территориальная изменчивость границ ареала составляет сотни километров, так что выразима в самом мелком масштабе. Для менее подвижных животных и растений такие колебания границ ареала невозможны, но в годы депрессий численности экологическая роль видов, находящихся в минимуме, практически равна нулю или стремится к нему. Исследователь может вид не обнаружить по чисто техническим причинам.
Возможности сопряжения и прогнозирования границ ареалов связаны с конгруэнтностью видов в составе биоценозов и сообществ (разд. 3.8.). Как показывает опыт, имеется соответствие видов и ценозов (см. ниже), и эти физиономически ясные черты позволяют отображать такие закономерности на картах. Однако довольно трудно (хотя и возможно) найти два вида, которые бы имели абсолютно сходное распределение в пространстве. Эту особенность Л. Г. Раменский (1924) и Г. А. Глизон (1926) сформулировали в виде индивидуалистической гипотезы, правила или принципа экологической индивидуальности видов: каждый вид пространственно распределен в соответствии со своими генетическими, физиологическими и другими биологическими особенностями и глубоко специфично относится к факторам среды, в том числе к другим видам. Как нет абсолютно идентичных особей (кроме однояйцевых близнецов, тем не менее нередко фенотипически отличающихся друг от друга), так нет и биоэкологически идентичных видов. Следовательно, их распространение сугубо индивидуально, а потому подвержено столь же индивидуальным нормам реакции на возникающие изменения среды жизни. Как сказано выше, этот принцип не абсолютен.
В виде регистрации факта можно сформулировать правило колебания границ ареала: многие виды не имеют четко отграниченной постоянной области распространения и временами либо исчезают на границе ареала, либо достигают исчезающе малой численности. В. Тишлер в 1955 г. сформулировал принцип воздействия факторов, согласно которому границы, размер и характер ареала вида или местообитания популяции обусловлены их биологическими особенностями, и наоборот, биологические особенности вида или популяции могут указывать (быть индикаторами) места, где их можно найти*.
| * Очень примитивно говоря, рыбы живут в воде — одна в реке, другая в озере, третья в море; одна на глубине, другая на поверхности и т. д.
| Точнее говоря, каждый вид или популяция входят в какие-то биоценозы и соответствующие экосистемы (биогеоценозы) и там, где есть вид или его специфическая популяция, очевидно, существует тот ценоз, куда они входят и где вырабатывают свою экологическую нишу. Если не существует такого соответствия, то, очевидно, либо вид, представленный популяцией, «не тот» (популяция имеет иную экологическую нишу), либо ценоз при физиономическом сходстве все же другой (иначе сформирован исторически и функционально).
| Ландшафтные зоны
| Основные типы местообитаний
| Характер размещения поселений
| | Тундра и лесотундра
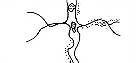
| 
| Долинно-речные (пойменно-речные, прибрежно-речные, речные острова)
| Ленточный
| | Тайга
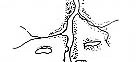
| 
| | Лесостепь
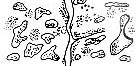
| 
| Болотные (заболоченные колки, осоково-кочкарниковые болота), озерные (прибрежные, займищные, сплавинные), лугово-полевые (поля, луга, огороды), долинно-речные (пойменные, прибрежные)
| Диффузный (междуречья) и ленточный (долины рек)
| | Степь
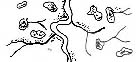
| 
| Озерные (займищно-озерные, озерно-сплавинные, прибрежно-озерные), долинно-речные (пойменные, прибрежные, займищные)
| Островной (междуречья) и ленточный (долины рек)
| | Предгорья
| 
| Долинно-ручьевые (в предгорьях: берега ручьев, болота в долинах ручьев; в горах: долины ручьев, межгорные котловины, субальпийские луга)
| Ленточный
| | Горы
| 
| Рис. 3.5. Структура поселений и особенности стационарного размещения водяной крысы в различных ландшафтных зонах Западной Сибири (рис. из кн.: Максимов А. А. Водяная крыса и борьба с ней в Западной Сибири. Новосибирск: 1959. С. 48)
| Как показывают широкомасштабные исследования в Сибири и на Дальнем Востоке, виды очень четко соответствуют ценозам, а если наблюдается разница в постоянно регистрируемом видовом составе всего в один вид, то ценоз явно резко отличается от географически или топографически соседних.
Как правило, смена видового состава происходит не на один, а на группу видов, и действительно, вид служит индикатором того сообщества, куда он входит.
Там, где природа сильно нарушена человеческой деятельностью, описанное выше правило соответствия вида и ценоза заметно не столь четко. Лоскутные природные образования складываются не в рамках бывших границ элементарных биогеоценозов, а составляют объединенные экосистемы топографически (но не экологически) более высокого иерархического уровня. Вид как бы вписывается не в «свой» изначальный ценоз, а в мозаику биогеоценотических парцелл разных экосистем, его экологическая ниша оказывается соотнесенной с этой мозаикой. Отсюда следует вывод, что правило соответствия вида и ценоза в этом случае распространяется на более географически широкие образования, а колебания границ ареалов могут в этом случае быть менее выраженными и не столь заметными.
Необходимо обратить внимание на то, что в ходе упомянутых процессов нарушается принцип территориальной общности физико-географических единиц (разд. 3.7.3), они укрупняются из-за мозаичности возникающих «обрывков» экосистем и их возрастающей отдаленности друг от друга. Иногда «промежуточные» экосистемы (по иерархии соподчинения) вообще исчезают, как, впрочем, и элементарные, в своем первозданном виде. Иерархия экосистем резко упрощается по числу уровней — сохраняется как целое лишь 2-й снизу уровень биома или ландшафтной полосы (см. главу 2) и только им соответствуют по распространению виды, занимая микроучастки, едва ли составляющие полноценные экосистемы (скорее, их парацеллы).
Там, где сохранилась первичная структура природных образований, соответствие вида и ценоза дает очень четкую пространственную картину кружева ареала, закономерно меняющегося при движении в географическом пространстве. Эта закономерность в разд. 3.6 была названа правилом топографического, или популяционного, кружева ареала. Это правило было сформулировано для топографического пространства одной географической точки. Теперь следует обратить внимание на изменения этого кружева в более широком пространстве — правило географической изменчивости кружева ареала. Обычно оно состоит в том, что на севере вид приурочен к приречным, низменным участкам, затем к югу заселяет более широкие пространства, в горах обитает лишь в локальных участках, а на южном пределе распространения вновь приурочен к приречным более влажным и прохладным местообитаниям. У каждого широко распространенного вида характер изменения кружева ареала специфичен, но смены происходят закономерно. Такие смены, пожалуй, лучше всего известны для водяной крысы в Сибири* (рис. 3.5). Пользуясь выявленными закономерностями, можно получить более надежные критерии для экстраполяции границ ареалов видов.
| * Очень показательная схема была опубликована (с. 48) в сборнике «Водяная крыса и борьба с ней в Западной Сибири». Новосибирское кн. изд-во, 1959 (Максимов А. А. Распределение водяной крысы по зонам и ландшафтам Западной Сибири. С. 19—48). См. также Пантелеев П А Популяционная экология водяной полевки и меры борьбы. М.: Наука, 1968. 255 с. ** РеймерсН. Ф. Птицы и млекопитающие южной тайги Средней Сибири. М.-Л.: Наука, 1966. 420 с.; Атлас Сахалинской области. М.: ГУГК, 1967.
| Проверка приведенных теоретических положений в свое время была сделана автором на Дальнем Востоке методом артиллерийской «вилки», т. е. были намечены точки, где вид уже не наблюдался и где наверняка существовал, а в промежуточном регионе по характеру растительности части экосистемы должен был бы присутствовать. В этой срединной точке проводились исследования. Все ожидавшиеся виды (в том числе не склонные к флуктуации границ ареала) были обнаружены вне зависимости от того, на каком расстоянии от точки наблюдения были проведены на картах ранее известные предельные границы их распространения. Часто находка была в сотнях километров к северу от «северного предела распространения» вида.
Правило соответствия вида и ценоза позволило автору предложить новый тип зоогеографических и шире — биогеографических — карт**, основанных на экосистемном подходе. При этом правило колебаний границ ареала в мелком масштабе удалось отразить лишь частично. Зато в крупном масштабе оно вполне может быть реализовано — ценозы, где наблюдается пульсация ареалов, графически на картах отличаются от тех, где вид постоянно присутствует.
Наиболее общим объяснением причин формирования границ ареала вида служит правило ограничивающих факторов: факторы среды, наиболее удаляющиеся от оптимума экологических потребностей вида, лимитируют возможности его существования в данных условиях. Поскольку к лимитирующим факторам относятся любые условия существования вида — как абиотические, так и биотические, включая антропогенные,— правило ограничивающих факторов, ведущее свое начало от группы законов минимума (см. разд. 3.5.2), включая закон толерантности Шелфорда (см. разд. 3.5.1), практически дополнительно ничего не объясняет, а лишь резюмирует перечисленные закономерности.
Для объяснения механизма действия принципа воздействия факторов В. Тишлера (см. выше) было предложено несколько теоретических оснований. Одно из них — правило стимулирующего действия температур В. Шелфорда — Т. Парка, в умеренных широтах определяющее стимулирующее воздействие, которое оказывают изменения температуры среды обитания (правило сформулировано независимо друг от друга В. Э. Шел-фордом в 1929 г. и Т. Парком в 1930 г.). Это правило в 1972 г. А. Н. Голиков и О. А. Скарлато дополнили правилом лимитирующего значения крайних летних и зимних температур, или «биогеографическим методом» указанных авторов: в умеренных широтах Северного полушария распространение видов ограничено необходимыми для размножения температурами. Эта потребность является генотипически закрепленным видовым признаком и прямо не связана с диапазоном температур выживаемости особей вида. У видов, переносящих широкие колебания температур, температура размножения часто более контрастна, чем у видов с относительно низким порогом выносливости к переменам количества тепла.
Очевидно, нельзя считать, что температура — абсолютно определяющий фактор в формировании границ ареала. Существенную роль играют влажность, субстраты, межвидовые отношения, преследование человеком, а в водной среде также скорость движения ее потоков и соленость вод. Саму температуру регулируют складывающийся мезо-, микро- и биоклимат. Играют роль и генетические закономерности взаимоотношения видов.
Среди них выделяется правило викариата Д. Джордана: ареалы близкородственных форм организмов (правило установлено для животных) обычно занимают смежные территории и существенно не перекрываются; родственные формы, как правило, викарируют, т. е. географически замещают друг друга. Согласно правилу представительства рода одним видом А. Монара (1919 г.), в однородных условиях и на ограниченной территории систематический (таксономический) род, как правило, представлен только одним видом. Видимо, это связано с близостью экологических ниш видов одного рода.
В связи с этим необходимо вспомнить группу закономерностей, связанных с принципом конкурентного исключения, или законом (теоремой) Г. Ф. Гаузе (1934 г.): два вида с близкими экологическими требованиями длительное время не могут занимать одну экологическую нишу и, как правило, входить в одну экосистему. Следовательно, одновременно происходит и географическое замещение видов, что констатирует принцип видо-родового представительства Й. Иллиеса: поскольку два близкородственных вида не могут занимать одинаковые экологические ниши в одном биотопе, и, соответственно, в биоценозе, богатые видами роды обычно представлены в экосистеме единственным своим представителем. Математически подобные отношения были доказаны уравнениями А. Лотки — В. Вольтерры (независимо этими авторами в 1925 и 1926 гг.). Однако поскольку принцип конкурентного исключения, видимо, не абсолютно универсален, был сформулирован принцип сосуществования, или парадокс Дж. Хатчинсона: два вида в порядке исключения могут сосуществовать в одной экологической нише, и если близкие виды вынуждены использовать одни и те же ресурсы, то естественный отбор может благоприятствовать их сосуществованию, и они способны эволюционировать в близком направлении. Парадокс Дж. Хатчинсона наблюдается главным образом в водных экосистемах*.
| * Кроме того, особи морфологически сходных видов, обитающих совместно, статистически различаются по размерам.
| Эмпирическим обобщением всех биогеографических закономерностей, касающихся ареалов и условий существования видов внутри них, служит правило географического оптимума: условия обитания вида наиболее оптимальны для него в центре ареала. Очевидно, предполагается, что абиотическая и биотическая среда тут наиболее благоприятна, а экологическая ниша выработана с наибольшим совершенством. Однако часто имеется в виду не пространственно-геометрический центр ареала, а его экологический «центр». Например, для морских котиков, многих других морских млекопитающих, колониальных птиц и любых наземных видов с дизъюнктивным или другим типом прерывистого ареала едва ли геометрический центр их ареала будет оптимален для жизни. Вид может вообще тут отсутствовать. Скорее, понятийно наоборот, экологический оптимум формирует часть ареала с наибольшей плотностью видового населения — оно имеет наивысшую плотность и общую численность в наиболее благоприятных местах существования популяций вида. Эти места могут располагаться как в глубине очерченного ареала, так и у его географических границ. В «чистом» виде правило географического оптимума справедливо лишь для видов с обширным сплошным ареалом, как правило занимающим всю либо большую часть биома или географической зоны. Однако и в этом случае нередко возможны варианты, когда наиболее благоприятными оказываются какие-то края области, распространения вида. Например, многие таежные животные (лось, волк и др.) явно приурочены не к глухой срединной тайге, а к так называемой подтаежной южной ее полосе. Здесь больше кормов по естественным причинам и из-за нарушения лесов человеком, к тому же теплее, и как правило, менее глубок снежный покров.
Ясно, что перечисленные биогеографические закономерности неравнозначны по глубине обобщения и вкладываемому смыслу.
Совершенно очевидно, что все закономерности системы «организм — среда» (разд. 3.5) отражаются на формировании границ ареалов видов. Наибольшее индикаторное значение имеет правило соответствия вида и ценоза, которое хотя и не объясняет причин такого соответствия, но отражает реально существующий факт, возникающий на почве сложного переплетения многих закономерностей генетического, аутоэкологического и синэкологического характера. Они формируют кружево видового ареала. Его географические пределы зависят от толерантности особей вида к физическим условиям среды и способности выработать экологическую нишу в одном или в нескольких ценозах, само существование которых определяется свойствами не столько отдельных видов, сколько их сообществ (разд. 3.8). Однако и тут важны характеристики видов-средообразователей — эдификаторов среды. Лишь во взаимодействии свойств популяций и их системных объединений складывается картина ареала. Поэтому все эти закономерности важны для анализа его границ и особенностей.
Нельзя сказать, чтобы внутренние механизмы формирования ареалов были до конца ясны, а главное, складывались в совокупность знаний, вполне пригодную для практической деятельности. Лучше обстоит дело с описанием внешних свойств географически сменяющих друг друга популяций, особенно животных. Эти закономерности сведены в следующем разделе.
|




