Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Глава 2. Морфология и классификация микробовСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
2.1. СИСТЕМАТИКА И НОМЕНКЛАТУРА МИКРОБОВ Микробы представлены доклеточными (вирусами - царство Virae) и клеточными (бактериями, архебактериями, грибами и простейшими) формами жизни. Клеточные формы жизни представлены прокариотами (бактериями), которые не имеют оформленного ядра и организованных органелл, и эукариотами (грибами и простейшими), клетки которых имеют ядро с ядерной оболочкой и ядрышком, а цитоплазма состоит из высокоорганизованных органелл (митохондрии, аппарат Гольджи и др.). Клеточные формы называют микроорганизмами. Различают следующие таксономические категории микроорганизмов: домен, царство, отдел, класс, порядок, семейство, род, вид, подвид и др. Царства микроорганизмов объединены в три домена (или империи) - Bacteria, Archaea и Eukarya: — домен Bacteria включает прокариоты, являющиеся настоящими бактериями (эубактериями); — домен Archaea включает прокариоты, являющиеся археями, или архебактериями; — домен Eukarya включает царства Protozoa (простейших), Eumycota (настоящих грибов) и Chromista (хромовиков) - новое царство, образованное в результате реклассификации некоторых простейших и грибов из более раннего устаревшего таксона - царства грибов (Fungi, Mycota). Одна из основных таксономических категорий - вид (species), т.е. совокупность особей, объединенных по близким свойствам, но отличающихся от других представителей рода. Совокупность однородных микроорганизмов, выделенных на питательной среде, характеризуемых сходными морфологическими, тинкториальными (отношением к красителям) и другими свойствами, называется чистой культурой. Чистая культура микроорганизмов, выделенных из определенного источника и отличающихся от других представителей вида, называется штаммом. Близким к понятию штамма является понятие клона. Для обозначения некоторых совокупностей микроорганизмов, отличающихся по тем или иным свойствам, употребляется суффикс -var- (разновидность) вместо ранее применяемого -type-. Именно поэтому микроорганизмы в зависимости от характера различий обозначают как морфовары (отличие по морфологии), резистентовары (отличие по устойчивости, например, к антибиотикам), серовары (отличие по антигенам), фаговары (отличие по чувствительности к бактериофагам), биовары (отличие по биологическим свойствам) и т. д. 2.2. КЛАССИФИКАЦИЯ И МОРФОЛОГИЯ БАКТЕРИЙ Бактерии относятся к прокариотам, т.е. доядерным организмам, поскольку у них имеется примитивное ядро без оболочки, ядрышка, гистонов, а в цитоплазме отсутствуют высокоорганизованные органеллы (митохондрии, аппарат Гольджи, лизосомы и др.). Название вида бактерий соответствует бинарной номенклатуре, т.е. состоит из двух слов. Например, возбудитель сифилиса пишется как Treponema pallidum. Первое слово - название рода и пишется с прописной буквы, второе слово обозначает вид и пишется со строчной буквы. При повторном упоминании вида родовое название сокращается до начальной буквы, например: T. pallidum. Согласно «Руководству» Берджи последних изданий (2001-2010), бактерии делят на два домена: Bacteria и Archaea (табл. 2.1). Подразделение бактерий (в домене Bacteria) по особенностям строения клеточной стенки связано с вариабельностью их окраски в тот или иной цвет по методу Грама. По этому методу, предложенному в 1884 г. датским ученым Х. Грамом, бактерии делятся на грамположительные, окрашиваемые в сине-фиолетовый цвет, и грамотрицательные, красящиеся в красный цвет. Таблица 2.1. Характеристика доменов Bacteria и Archaea
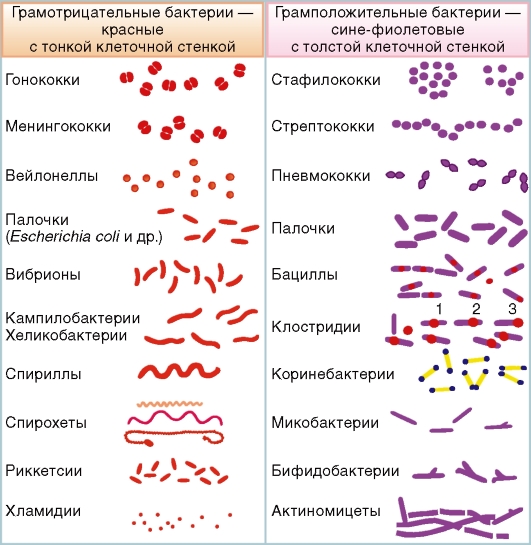
Грамотрицательные бактерии имеют тонкую клеточную стенку (рис. 2.1). К ним относятся сферические (кокки: гонококки, менингококки, вейлонеллы), извитые (спирохеты и спириллы), а также палочковидные и наиболее мелкие бактерии (риккетсии и хламидии - облигатные внутриклеточные паразиты). Грамположительные бактерии имеют толстую клеточную стенку. К ним относятся сферические (кокки: стафилококки, стрептококки, пневмококки), палочковидные и ветвящиеся нитевидные формы (актиномицеты). 2.2.1. ФОРМЫ БАКТЕРИЙ Бактерии имеют форму кокков, палочек, спиралей и другие формы (см. рис. 2.1). Кокки - шаровидные бактерии размером 0,5-1,0 мкм; по их взаимному расположению различают микрококки, диплококки, стрептококки, тетракокки, сарцины и стафилококки. — Микрококки (от греч. micros - «малый») - отдельно расположенные клетки. — Диплококки (от греч. diploos - «двойной») расположены парами (пневмококк, гонококк, менингококк), так как клетки после деления не расходятся. Пневмококк (возбудитель пневмонии) имеет с противоположных сторон ланцетовидную форму, а гонококк (возбудитель гонореи) и менингококк (возбудитель эпидемического менингита) имеют форму кофейных зерен, обращенных вогнутой поверхностью друг к другу (см. рис. 2.1).
Рис. 2.1. Морфологические и тинкториальные свойства бактерий. Некоторые бактерии образуют споры, расположенные центрально (1), субтерминально (2) или терминально (3) — Стрептококки (от греч. streptos - «цепочка») - клетки округлой формы, составляющие цепочку вследствие деления клеток в одной плоскости. — Сарцины (от лат. sarcina - «связка, тюк») расположены в виде пакетов из 8 кокков и более, так как они образуются при делении клетки в трех взаимно перпендикулярных плоскостях. — Стафилококки (от греч. staphyle - «виноградная гроздь») - кокки, расположенные в виде грозди винограда в результате деления в разных плоскостях. Палочковидные бактерии различаются по размерам, форме концов клетки и взаимному расположению клеток. Длина клеток варьирует от 1 до 10 мкм, толщина - от 0,5 до 2 мкм. Концы палочек могут быть как бы обрезанными (сибиреязвенная бацилла), закругленными (кишечная палочка), заостренными (фузобактерии) или в виде утолщения. В последнем случае палочка похожа на булаву (коринебактерии дифтерии). Слегка изогнутые палочки называются вибрионами (холерный вибрион). Большинство палочковидных бактерий расположены беспорядочно. Если после деления клетки остаются связанными общими фрагментами клеточной стенки и не расходятся, то они располагаются под углом друг к другу (коринебактерии дифтерии) или образуют цепочку (сибиреязвенная бацилла). Извитые формы - спиралевидные бактерии, к которым относятся кампилобактерии, хеликобактерии, спириллы и спирохеты. Спириллы имеют вид штопорообразно извитых клеток. Спирохеты - тонкие, длинные, извитые (спиралевидной формы) бактерии, отличающиеся от спирилл подвижностью, обусловленной сгибательными изменениями клеток. Они имеют тонкую наружную мембрану клеточной стенки, окружающую протоплазматический цилиндр с цитоплазматической мембраной, и периплазматические фибриллы (жгутики). Фибриллы участвуют в передвижении спирохет, придавая клеткам вращательное, сгибательное и поступательное движение. Спирохеты представлены тремя родами, патогенными для человека: Treponema, Borrelia, Leptospira. — Трепонемы (род Тreропета) имеют вид тонких, штопорообразно закрученных нитей с 8-12 равномерными мелкими завитками, например, T. pallidum - возбудитель сифилиса. — Боррелии (род Borrelia) имеют по 3-8 крупных завитков. К ним относятся возбудитель возвратного тифа (B. recurrentis) и возбудители болезни Лайма (B. burgdorferi и др.). — Лептоспиры (род Leptospira) имеют неглубокие и частые завитки в виде закрученной веревки. Концы этих спирохет изогнуты наподобие крючков с утолщениями на концах. Образуя вторичные завитки, они приобретают вид букв С или S (например, L. interrogans - возбудитель лептоспироза). Ветвящиеся нитевидные формы и палочки неправильной формы представлены актиномицетами и родственными с ними бактериями (нокардиями, микобактериями, коринебактериями и др.). Актиномицеты - ветвящиеся нитевидные или палочковидные грамположительные бактерии. Свое название (от греч. actis - «луч» и mykes - «гриб») они получили в связи с образованием в пораженных тканях друз - гранул из плотно переплетенных нитей в виде лучей, отходящих от центра и заканчивающихся колбовидными утолщениями. С актиномицетами сходны так называемые нокардиоподобные (нокардиоформные) актиномицеты -палочковидные, неправильной формы. К ним относят бактерии родов Corynebacterium, Mycobacterium, Nocardia и др. Нокардиоподобные актиномицеты отличаются наличием в клеточной стенке сахаров - арабинозы, галактозы, а также миколовых кислот и больших количеств жирных кислот. Миколовые кислоты и липиды клеточных стенок обусловливают кислотоустойчивость бактерий, в частности, микобактерий туберкулеза и лепры (при окраске по Цилю-Нильсену они имеют красный цвет, а некислотоустойчивые бактерии и элементы ткани, мокроты - синий цвет). Риккетсии - мелкие грамотрицательные палочковидные бактерии (0,3-2,0 мкм), облигатные (обязательные) внутриклеточные паразиты, рамножающиеся в цитоплазме, а некоторые - в ядре инфицированных клеток. Обитают в членистоногих (вшах, блохах, клещах), которые являются их хозяевами или перенос чиками. Свое название риккетсии получили по имени Х. Т. Риккетса - американского ученого, впервые описавшего одного из возбудителей. У человека риккетсии вызывают эпидемический сыпной тиф (Rickettsia prowazekii), клещевой риккетсиоз (R. sibirica) и другие риккетсиозы. Хламидии - облигатные внутриклеточные кокковидные грамотрицательные бактерии, размножающиеся только в живых клетках. Вне клеток хламидии имеют сферическую форму (0,3 мкм), метаболически неактивны и называются элементарными тельцами. Элементарные тельца, попав в клетку, увеличиваются и превращаются в делящиеся ретикулярные тельца, образуя скопления (включения) в вакуолях. Из ретикулярных телец образуются элементарные тельца, которые выходят из клеток и вступают в новый цикл, инфицируя другие клетки. У человека хламидии вызывают поражения глаз (трахому, конъюнктивит), урогенитальной системы, легких и др. Микоплазмы - мелкие бактерии (0,3-0,8 мкм), не имеющие клеточной стенки; окружены только цитоплазматической мембраной, содержащей стеролы. Вследствие отсутствия клеточной стенки микоплазмы осмотически чувствительны. Имеют кокковидную, нитевидную или колбовидную формы. Микоплазмы вызывают у человека атипичную пневмонию (Mycoplasma pneumoniae) и поражение мочеполовой системы (M. hominis и др.). 2.2.2. СТРУКТУРА БАКТЕРИАЛЬНОЙ КЛЕТКИ Бактерии измеряют в микрометрах (мкм), а их структурные компоненты - в нанометрах (нм); один микрометр равен 1000 нм. Вследствие малых размеров структуру бактерий изучают с помощью электронной микроскопии целых клеток и их ультратонких срезов. Бактериальную клетку окружают клеточная стенка и цитоплазматическая мембрана, под которой находятся цитоплазма с включениями и примитивное ядро, называемое нуклеоидом. Имеются также дополнительные структуры: капсула, микрокапсула, слизь, жгутики, пили (рис. 2.2). Некоторые бактерии в неблагоприятных условиях образуют споры.
Рис. 2.2. Строение бактериальной клетки Клеточная стенка - прочная, упругая структура, придающая бактерии определенную форму и вместе сцитоплазматической мембраной (ЦПМ) сдерживающая высокое осмотическое давление в бактериальной клетке. В клеточной стенке грамположительных бактерий содержится небольшое количество полисахаридов, липидов, белков. Основной компонент клеточной стенки этих бактерий - многослойный пептидогликан, с которым ковалентно связаны тейхоевые кислоты (от греч. teichos - «стенка»). Имеются также липотейхоевые кислоты, которые гидрофобно связаны с ЦПМ (рис. 2.3). Эти полимеры участвуют в регуляции деления бактерии и в адгезии на клетках организма при инфицировании. Форму и прочность бактериям придает жесткая волокнистая структура многослойного пептидогликана.
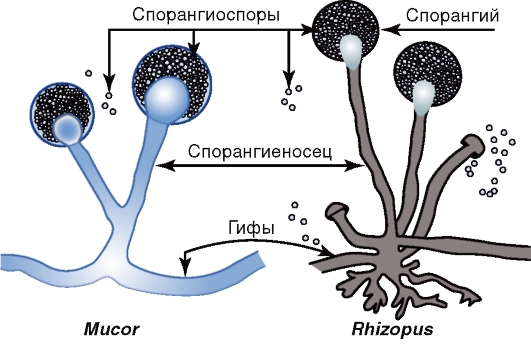
Рис. 2.3. Строение оболочек (клеточной стенки и цитоплазматической мембраны) грамположительных и грамотрицательных бактерий Пептидогликан представлен параллельно расположенными молекулами гликана, состоящего из повторяющихся остатков N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных гликозидной связью. Эти связи разрывает лизоцим, подавляя синтез клеточной стенки. Гликановые молекулы соединены поперечной пептидной связью из четырех аминокислот (например, L-аланин- D-глутаминовая кислота-L-лизин-D-аланин). Отсюда и название этого полимера - пептидогликан. Основу пептидной связи пептидогликана грамотрицательных бактерий составляют тетрапептиды, состоящие из чередующихся L- и D-аминокислот (например, L-аланин-D-глутаминовая кислота-мезодиаминопимелиновая кислота-D-аланин). Наличие элементов гликана (ацетилглюкозамина и ацетилмурамовой кислоты) и аминокислот тетрапептида (мезодиаминопимелиновой и D-глутаминовой кислот, D-аланина) - отличительная особенность бактерий, поскольку они отсутствуют у животных и человека. При окраске по Граму многослойный пептидогликан грамположительных бактерий после обработки мазка спиртом удерживает комплекс генцианового фиолетового с йодом, чем обусловлена синефиолетовая окраска клеток. Грамотрицательные бактерии, наоборот, после воздействия спиртом утрачивают краситель, что обусловлено меньшим количеством пептидогликана (5-10% массы клеточной стенки); они обесцвечиваются спиртом и после обработки фуксином или сафранином приобретают красный цвет. В состав клеточной стенки грамотрицательных бактерий входит наружная мембрана, связанная посредством липопротеина с подлежащим слоем пептидогликана (см. рис. 2.3). Наружная мембрана при электронной микроскопии ультратонких срезов бактерий имеет вид волнообразной трехслойной структуры, сходной с внутренней мембраной, которую называют цитоплазматической. Основной компонент этих мембран - двойной слой липидов. Наружная мембрана - мозаичная структура, представленная липополисахаридами, фосфолипидами и белками. Внутренний слой ее содержит фосфолипиды, а в наружном слое расположен липополисахарид (ЛПС). ЛПС наружной мембраны состоит из трех фрагментов: — липида А - консервативной структуры, практически одинаковой у грамотрицательных бактерий; — ядра, или стержневой, коровой части (от лат. core - «ядро»), относительно консервативного олигосахарида; — О-специфической цепи полисахарида, образованной повторяющимися идентичными олигосахаридными последовательностями. ЛПС заякорен в наружной мембране липидом А, обусловливающим токсичность ЛПС и отождествляемым поэтому с эндотоксином. Разрушение бактерий антибиотиками приводит к высвобождению большого количества эндотоксина, что может вызвать у больного эндотоксический шок. От липида А отходит ядро, или стержневая часть, ЛПС. O-специфическая цепь, отходящая от стержневой части молекулы ЛПС, обусловливает серогруппу, серо- вар (разновидность бактерий, выявляемую с помощью иммунной сыворотки) определенного штамма бактерий. Таким образом, с понятием ЛПС связаны представления об О-антигене, по которому можно дифференцировать бактерии. Бактерии с полноценным ЛПС образуют гладкие, с блестящей поверхностью колонии, получившие название S-форм (от англ. smooth - «гладкий»). Генетические изменения могут привести к дефектам, укорочению ЛПС бактерий и появлению в результате этого шероховатых колоний R-форм (от англ. rough - «шероховатый»). Белки наружной мембраны, называемые поринами, окаймляют гидрофильные поры, через которые проходятвода и мелкие гидрофильные молекулы. Между наружной мембраной и ЦПМ находится периплазматическое пространство, или периплазма, содержащая ферменты и компоненты транспортных систем. При нарушении синтеза клеточной стенки бактерий под влиянием лизоцима, пенициллина, защитных факторов организма и других соединений образуются клетки с измененной (часто шаровидной) формой: протопласты - бактерии, полностью лишенные клеточной стенки; сферопласты - бактерии с частично сохранившейся клеточной стенкой. Такие формы, утратившие способность к синтезу пептидогликана под влиянием антибиотиков или других факторов и способные размножаться, называются L-формами (от названия института им. Д. Листера, где они впервые были изучены). L-формы могут возникать и в результате мутаций. L-формы - осмотически чувствительные шаровидные или колбовидные клетки различной величины, в том числе и проходящие через бактериальные фильтры. Некоторые L-формы (нестабильные) при удалении фактора, приведшего к изменениям бактерий, могут реверсировать, возвращаясь в исходную бактериальную клетку. L-формы могут образовываться многими возбудителями инфекционных болезней. ЦПМ окружает наружную часть цитоплазмы бактерий и участвует в регуляции осмотического давления, транспорте веществ и энергетическом метаболизме клетки. По структуре она схожа с плазмалеммой клеток животных и состоит из двойного слоя липидов, главным образом фосфолипидов, с внедренными белками. Некоторые из них являются пермеазами, участвующими в транспорте веществ. При избыточном росте ЦПМ образует инвагинаты - впячивания в виде сложно закрученных мембранных структур, называемых мезосомами. Предполагают, что производные ЦПМ участвуют в делении клетки, обеспечивая энергией синтез клеточной стенки, в секреции веществ, спорообразовании, т.е. в процессах с высокой затратой энергии. Цитоплазма занимает основной объем бактериальной клетки и состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких гранул - рибосом, ответственных за синтез (трансляцию) белков. Рибосомы бактерий имеют размер около 20 нм и коэффициент седиментации 70S, в отличие от 80S-рибосом, характерных для эукариотических клеток. Именно поэтому некоторые антибиотики, связываясь с рибосомами бактерий, подавляют синтез бактериального белка, не влияя на синтез белка эукариотических клеток. В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, β-оксимасляной кислоты и полифосфатов (волютина). Характерное расположение гранул волютина выявляется у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки. Нуклеоид - эквивалент ядра у бактерий. Ядро бактерий, в отличие от эукариот, не имеет ядерной оболочки, ядрышка и основных белков (гистонов). У большинства бактерий в клетке содержится одна хромосома, представленная замкнутой в кольцо молекулой дезоксирибонуклеиновой кислоты - ДНК (иногда линейной ДНК). Некоторые бактерии имеют две хромосомы и более. Кроме нуклеоида, в бактериальной клетке имеются внехромосомные факторы наследственности - плазмиды - ковалентно замкнутые кольца ДНК. Плазмиды придают бактериям дополнительные свойства: устойчивость к антибиотикам (R-плазмиды), способность к передаче генетического материала при конъюгации (F-плазмиды) и др. Капсула - слизистая структура толщиной более 0,2 мкм, прочно связанная с клеточной стенкой бактерий и имеющая четко очерченные внешние границы. Капсула различима в мазках-отпечатках из патологического материала. В чистых культурах бактерий капсула образуется реже. Она выявляется при специальных методах окраски мазка по Бури-Гинсу, создающих негативное контрастирование веществ капсулы: тушь образует темный фон вокруг капсулы. Капсула состоит из полисахаридов (экзополисахаридов), иногда из полипептидов; например, у сибиреязвенной бациллы она состоит из полимеров D-глутаминовой кислоты и препятствует фагоцитозу бактерий. Многие бактерии образуют микрокапсулу - слизистое образование толщиной менее 0,2 мкм, выявляемое лишь при электронной микроскопии. От капсулы следует отличать слизь - мукоидные экзополисахариды, не имеющие четких внешних границ. Слизь растворима в воде. Жгутики бактерий - тонкие нити, берущие начало от ЦПМ, имеют большую длину, чем сама клетка, определяют подвижность бактериальной клетки. Жгутики состоят из белка - флагеллина (от. лат. flagellum -«жгутик»), являющегося антигеном (АГ) - так называемым Н-антигеном. Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен жгутиков, отходящих по периметру бактерии (перитрих), у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки. Пили (фимбрии, ворсинки) - нитевидные образования, более короткие и тонкие (3-10 нм × 0,3-10 мкм), чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина. Различают пили, ответственные за адгезию, т.е. за прикрепление бактерий к поражаемой клетке, а также пили, ответственные за питание, водно-солевой обмен и за передачу генетического материала, - половые F-пили, или конъюгационные пили. Споры - своеобразная форма покоящихся бактерий, образуемая при неблагоприятных условиях существования бактерий (высушивании, УФ-облучении, дефиците питательных веществ и др.). Внутри бактериальной клетки образуется одна спора - эндоспора. Образование спор способствует сохранению вида и не является способом размножения, как у грибов. Спорообразующие бактерии рода Bacillus, у которых размер споры не превышает диаметра клетки, называются бациллами. Спорообразующие бактерии (см. рис. 2.1), у которых размер споры превышает диаметр клетки, принимают форму веретена и называются клостридиями, например, бактерии рода Clostridium (от лат. dostridium - «веретено»). Споры кислотоустойчивы, поэтому окрашиваются по методам Ауески и Циля-Нильсена в красный цвет, а вегетативная клетка - в синий. Форма спор бывает овальной, шаровидной. Расположение спор в клетке может быть: — терминальным, т.е. на конце палочки (у возбудителя столбняка); — субтерминальным - ближе к концу палочки (у возбудителей ботулизма, газовой гангрены); — центральным (у сибиреязвенной бациллы). Процесс спорообразования (споруляция) проходит ряд стадий, в течение которых формируется многослойная, плохо проницаемая оболочка. Спора приобретает термоустойчивость, которую связывают с наличием в ней дипиколината кальция. Спора может долго сохраняться из-за наличия многослойной оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизма. В почве споры, например, возбудителей сибирской язвы и столбняка могут сохраняться десятки лет. В благоприятных условиях споры прорастают, проходя три последовательные стадии: активацию, инициацию, вырастание. При этом из одной споры образуется одна бактерия. 2.3. СТРОЕНИЕ И КЛАССИФИКАЦИЯ ГРИБОВ Грибы - многоили одноклеточные нефотосинтезирующие (бесхлорофильные) эукариотические микроорганизмы с толстой клеточной стенкой. Они имеют ядро с ядерной оболочкой, цитоплазму с органеллами, ЦПМ и многослойную ригидную клеточную стенку, состоящую из нескольких типов полисахаридов (маннанов, глюканов, целлюлозы, хитина), а также белка и липидов. Некоторые грибы образуют капсулу. ЦПМ содержит гликопротеины, фосфолипидыи эргостеролы (в отличие от холестерина - главного стерола тканей млекопитающих). Грибы широко распространены в природе, особенно в почве. Различают два основных типа грибов: гифальные и дрожжевые. Гифальные (плесневые) грибы образуют ветвящиеся тонкие нити (гифы), сплетающиеся в грибницу, или мицелий (плесень). Гифы низших грибов не имеют перегородок. Гифы высших грибов разделены перегородками, или септами, с отверстиями. Дрожжевые грибы (дрожжи) в основном имеют вид отдельных овальных клеток. Дрожжи - одноклеточные грибы. Они имеют половое и бесполое размножение. При бесполом размножении дрожжи образуют почки или делятся, что приводит к одноклеточному росту. Могут образовывать псевдогифы и ложный мицелий (псевдомицелий), состоящие из цепочек удлиненных клеток в виде сарделек. Грибы, аналогичные дрожжам, но не имеющие полового способа размножения, называют дрожжеподобными. Они размножаются только бесполым способом - почкованием или делением. Многие грибы характеризуются диморфизмом - способностью к гифальному (мицелиальному) или дрожжеподобному росту в зависимости от условий культивирования. Например, в инфицированном организме они растут в виде дрожжеподобных клеток (дрожжевая фаза), а на питательных средах образуют гифы и мицелий. Размножение грибов происходит половым и бесполым (вегетативным) способами. Половое размножение грибов происходит с образованием гамет, половых спор и других половых форм. Половые формы называются телеоморфами. Бесполое размножение грибов происходит с образованием соответствующих форм, называемых анаморфами. Такое размножение происходит почкованием, фрагментацией гиф и бесполыми спорами. Эндогенные споры (спорангиоспоры) созревают внутри округлой структуры - спорангия (рис. 2.4).
Рис. 2.4. Грибы родов Mucor и Rhizopus Экзогенные споры (конидии) формируются на кончиках плодоносящих гиф - так называемых конидиеносцах (рис. 2.5).
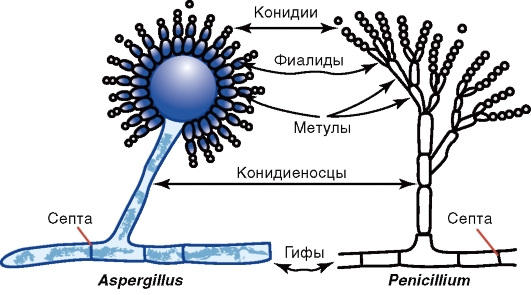
Рис. 2.5. Грибы родов Penicillium и Aspergillus Царство настоящих грибов Eumycota включает четыре типа (Phylum) грибов, имеющих медицинское значение: зигомицеты (Zygomycota), аскомицеты (Ascomycota), базидиомицеты (Basidiomycota) и дейтеромицеты (Deiteromycota). Различают совершенные и несовершенные грибы. Совершенные грибы имеют половой способ размножения; к ним относятся зигомицеты, аскомицеты и базидиомицеты. Несовершенные грибы имеют только бесполый способ размножения; к ним относятся дейтеромицеты. Зигомицеты относятся к низшим грибам. Они включают виды родов Мucor, Rhizopus и др. Распространены в почве, воздухе и способны вызывать зигомикоз (мукоромикоз) легких, головного мозга и других органов человека и животных. Половое размножение у зигомицетов осуществляется путем образования зигоспор. При бесполом размножении этих грибов на плодоносящей гифе - спорангиеносце - образуется спорангий с многочисленными спорангиоспорами (см. рис. 2.4). Аскомицеты имеют септированный мицелий (за исключением одноклеточных дрожжей). Свое название они получили от основного органа плодоношения - сумки, или аска, содержащего 4 или 8 гаплоидных половых спор (аскоспор). К аскомицетам относятся отдельные представители (телеоморфы) родов Aspergillus, Penicillium и др. Большинство грибов родов Aspergillus и Penicillium являются анаморфами, т.е. размножаются только бесполым путем и по этому признаку относятся к несовершенным грибам. Они отличаются особенностями формирования плодоносящих гиф (рис. 2.6).
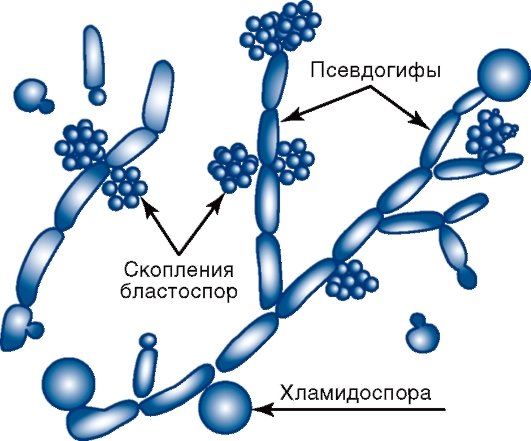
Рис. 2.6. Candida albicans У грибов рода Aspergillus (леечной плесени) на концах плодоносящих гиф (конидиеносцах) имеются утолщения - стеригмы, на которых образуются цепочки спор - конидии. Некоторые виды аспергилл могут вызывать аспергиллезы и афлатоксикозы. Плодоносящая гифа у грибов рода Penicillium (кистевика) напоминает кисточку, так как из нее (на конидиеносце) образуются утолщения, разветвляющиеся на более мелкие структуры - стеригмы, на которых находятся цепочки конидий. Пенициллы могут вызывать заболевания - пенициллиозы. Представителями аскомицетов являются также дрожжи - одноклеточные грибы овальной формы, диаметром 3-15 мкм. Заболевания, вызываемые некоторыми видами дрожжей, получили название дрожжевых микозов. Многие виды аскомицетов являются продуцентами антибиотиков, используемыми в биотехнологиях. Базидиомицеты - шляпочные съедобные и ядовитые грибы с септированным мицелием. Они образуют половые споры (базидиоспоры) путем отшнуровывания от базидия - концевой клетки мицелия, гомологичной аску. Дейтеромицеты (несовершенные грибы) являются условным, формальным типом грибов, который объединяет грибы, не имеющие полового способа размножения. Слово «формальный» означает, что потенциально эти грибы могут иметь половой способ размножения; при установлении этого факта грибы переносят в один из известных типов - Ascomycota или Basidiomycota и им присваивают название телеоморфной формы. Дейтеромицеты образуют септированный мицелий, размножаются только бесполым путем за счет формирования неполовых спор - конидий. К дейтеромицетам относятся несовершенные дрожжи (дрожжеподобные грибы), например, некоторые грибы рода Candida, поражающие кожу, слизистые оболочки и внутренние органы (кандидоз). Они имеют овальную форму (см. рис. 2.6), диаметр 2-5 мкм, делятся почкованием, образуют псевдогифы (псевдомицелий) в виде цепочек из удлиненных клеток и септированные гифы. Эти грибы называются дрожжеподобными, в отличие от истинных дрожжей, которые относятся к аскомицетам, образующим аскоспоры. Для Candida albicans характерно образование хламидоспор. 2.4. СТРОЕНИЕ И КЛАССИФИКАЦИЯ ПРОСТЕЙШИХ Простейшие - эукариотические одноклеточные микроорганизмы, составляющие царство Protozoa (от греч. protos - «первый», zoon - «животное») в домене Eukarya. Недавно выделено новое царство - Chromista (хромовиков), в которое вошли некоторые простейшие и грибы (бластоцисты, оомицеты, Rhinosporidium seeberi и др.). Простейшие имеют ядро с ядерной оболочкой и ядрышком, их цитоплазма состоит из эндоплазматического ретикулума, митохондрий, лизосом, многочисленных рибосом и др. Размеры простейших колеблются от 2 до 100 мкм. Снаружи они окружены мембраной (пелликулой) - аналогом ЦПМ клеток животных. Простейшие имеют органы движения (жгутики, реснички, псевдоподии), питания (пищеварительные вакуоли) и выделения (сократительные вакуоли). Жгутики отходят от блефаропласта. Питание простейших происходит путем фагоцитоза или образования особых структур, а размножение - бесполым (двойным или множественным делением - шизогония) или половым (спорогония) путями. При неблагоприятных условиях многие из них образуют цисты - покоящиеся стадии, устойчивые к изменению температуры, влажности и др. При окраске по Романовскому-Гимзе ядро простейших имеет красный цвет, а цитоплазма - синий. К царству Protozoa относятся амебы, жгутиконосцы, споровики и реснитчатые (рис. 2.7). — Амебы представлены Entamoeba histolytica - возбудителем амебиаза человека (амебной дизентерии), а также свободноживущими и непатогенными амебами (кишечной амебой и др.). Они размножаются бесполым путем (бинарно). Жизненный цикл амеб состоит из двух стадий: трофозоита (растущей подвижной клетки, малоустойчивой) и цисты. Трофозоиты передвигаются путем образования псевдоподий, с помощью которых происходят захват и погружение в цитоплазму клеток питательных веществ. Из трофозоита образуется циста, устойчивая к внешним факторам. Циста, попадая в кишечник, превращается в трофозоит. — Жгутиконосцы включают жгутиконосцев крови и других тканей: ◊ жгутиконосцев крови и тканей (лейшманий - возбудителей лейшманиозов; трипаносом - возбудителей африканского трипаносомоза, или сонной болезни, и болезни Шагаса); ◊ жгутиконосцев кишечника (лямблии - возбудителя лямблиоза); ◊ жгутиконосцев мочеполовой системы (трихомонаду влагалищную - возбудителя трихомоноза). Эти простейшие характеризуются наличием жгутиков, например, у лейшманий - один жгутик, у трихомонад - четыре свободных жгутика и один жгутик, соединенный с короткой ундулирующей мембраной. — Споровики представлены: ◊ кровяными паразитами, к которым относятся плазмодии малярии и бабезии - возбудители пироплазмоза; ◊ кишечными и тканевыми паразитами, к которым относятся токсоплазма - возбудитель токсоплазмоза, криптоспоридии - возбудители криптоспоридиоза, саркоцисты - возбудители саркоцистоза, изоспоры - возбудители изоспороза, циклоспоры - возбудители циклоспоридиоза. Паразиты имеют апикальный комплекс, который позволяет им проникнуть в клетку хозяина для последующего внутриклеточного паразитизма. Каждый из этих простейших имеет сложное строение и свои особенности жизненного цикла. Так, например, жизненный цикл возбудителя малярии характеризуется чередованием полового (в организме комаров Anopheles) и бесполого (в клетках печени и эритроцитах человека, где они размножаются множественным делением) размножения. - Реснитчатые представлены балантидиями, поражающими толстую кишку человека (балантидиазная дизентерия). Балантидии имеют стадии трофозоита и цисты. Трофозоит подвижен, имеет многочисленные реснички, более тонкие и короткие, чем жгутики.
Рис. 2.7. Морфология основных представителей простейших 2.5. СТРОЕНИЕ И КЛАССИФИКАЦИЯ ВИРУСОВ Вирусы относятся к царству Virae (от лат. virus - «яд»). Это мельчайшие микроорганизмы («фильтрующиеся агенты»), не имеющие клеточного строения, белоксинтезирующей системы, содержащие один тип нуклеиновой кислоты (или ДНК, или рибонуклеиновой кислоты - РНК). Вирусы как облигатные внутриклеточные паразиты размножаются в цитоплазме или ядре клетки. Они являются автономными генетическими структурами и отличаются особым, разобщенным (дизъюнктивным) способом размножения (репродукции): в клетке отдельно синтезируются нуклеиновые кислоты вирусов и их белки, а затем происходит их сборка в вирусные частицы. Сформированная вирусная частица называется вирионом. Морфологию и структуру вирусов изучают с помощью электронной микроскопии, так как их размеры малы и сравнимы с толщиной оболочки бактерий. Форма вирионов может быть палочковидной (вирус табачной мозаики), пулевидной (вирус бешенства), сферической (вирусы полиомиелита, вирус иммунодефицита человека - ВИЧ), нитевидной (филовирусы) или в виде сперматозоида (многие бактериофаги - см. главу 3). Наиболее мелкими являются парвовирусы (18 нм) и вирус полиомиелита (около 20 нм), наиболее крупным - вирус натуральной оспы (около 350 нм). Различают ДНК- и РНК-содержащие вирусы. Геном вирусов содержит от шести до нескольких сотен генов и представлен различными видами нуклеиновых кислот: дву-, однонитевыми, линейными, кольцевыми, фрагментированными. Среди однонитевых РНК-содержащих вирусов различают вирусы с плюс-нитью РНК и вирусы с минус-нитью РНК (полярность РНК). Плюс-нить РНК (позитивная нить) выполняет наследственную (геномную) функцию и функцию матричной, или информационной, РНК (иРНК), являясь матрицей для белкового синтеза на рибосомах инфицированной клетки. Плюс-нить РНК является инфекционной: при введении в чувствительные клетки она способна вызвать инфекционный процесс. Минус-нить (негативная нить) выполняет только наследственную функцию; для синтеза белка на минус-нити РНК синтезируется комплементарная ей нить. У некоторых вирусов РНК-геном содержит плюс- и минус-сегменты РНК. Геном вирусов способен включаться в геном клетки в виде провируса, проявляясь генетическим паразитом клетки. Нуклеиновые кислоты некоторых вирусов, например вирусов герпеса, могут находиться в цитоплазме инфицированных клеток, напоминая плазмиды. Различают простые вирусы (например, вирусы полиомиелита, гепатита А) и сложные вирусы (например, вирусы кори, гриппа, герпеса). Простые, или безоболочечные, вирусы (рис. 2.8) имеют
|
||||||||
|
|
Последнее изменение этой страницы: 2021-04-05; просмотров: 612; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.68.167 (0.015 с.) |