Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Глава 4. Физиология микроорганизмов
4.1. ОСОБЕННОСТИ ФИЗИОЛОГИИ БАКТЕРИЙ 4.1.1. ХИМИЧЕСКИЙ СОСТАВ БАКТЕРИАЛЬНОЙ КЛЕТКИ Бактериальная клетка на 80-90% состоит из воды, и только 10% приходится на долю сухого вещества. Вода в клетке находится в свободном или связанном состоянии. Она выполняет механическую роль в обеспечении тургора, участвует в гидролитических реакциях. Удаление воды из клетки путем высушивания приводит к приостановке процессов метаболизма, прекращению размножения, а для многих бактерий - к гибели. В то же время особый способ высушивания микроорганизмов в вакууме из замороженного состояния - лиофилизация, которая обеспечивает сохранение жизнеспособности большинства микроорганизмов. Лиофилизацию используют для приготовления проб, пригодных для длительного хранения, и при приготовлении вакцин. Состав сухого вещества бактерий распределен следующим образом: 52% - белки, 17% - углеводы, 9% - липиды, 16% - РНК, 3% - ДНК и 3% - минеральные вещества. Белки выполняют роль ферментов, а также входят в состав ЦПМ и ее производных, клеточной стенки, жгутиков, спор и некоторых капсул. Некоторые бактериальные белки являются АГ и токсинами бактерий. Углеводы представлены в бактериальной клетке в виде моно-, ди-, олиго- и полисахаридов. Они находятся в составе некоторых капсул, клеточной стенки. Крахмал и гликоген выполняют функцию запасных питательных веществ. Некоторые полисахариды принимают участие в формировании АГ. Липиды, или жиры, входят в состав ЦПМ и ее производных; клеточной стенки грамотрицательных бактерий, эндотоксина грамотрицательных бактерий. Они служат запасными веществами, а в составе ЛПС формируют АГ. У некоторых бактерий в клетке находятся воски, эфиры миколовой кислоты. Микоплазмы - единственные представители царства Procaryotae, имеющие в составе ЦПМ стеролы. Другие бактерии в составе ЦПМ и ее производных стеролов не имеют. Нуклеиновые кислоты. В бактериальной клетке присутствуют все типы РНК: иРНК, транспортная РНК (тРНК), рибосомная РНК (рРНК), двухцепочечная ДНК. Пуриновые и пиримидиновые нуклеотиды - строительные блоки, из которых построены нуклеиновые кислоты. Минеральные вещества представлены N, S, Р, Са, К, Mg, Fe, Mn, а также микроэлементами: Zn, Cu, Co, Ва. Они входят в структуру белков, нуклеотидов и выступают в роли активаторов ферментов.
4.1.2. КЛАССИФИКАЦИЯ БАКТЕРИЙ ПО ТИПАМ ПИТАНИЯ И СПОСОБАМ ПОЛУЧЕНИЯ ЭНЕРГИИ Основной целью метаболизма бактерий является рост, т.е. координированное увеличение всех компонентов клетки. В связи с тем что основными компонентами бактериальной клетки являются органические соединения, белки, углеводы, нуклеиновые кислоты и липиды, остов которых построен из атомов углерода, для роста требуется постоянный приток атомов углерода. В зависимости от источника усвояемого углерода бактерии подразделяют: — на аутотрофы (от греч. autos - «сам», trophe - «питание») - использующие для построения своих клеток неорганический углерод в виде СО2; — гетеротрофы (от греч. heteros - «другой») - использующие органический углерод. Легкоусвояемыми источниками органического углерода являются гексозы, многоатомные спирты, аминокислоты, липиды. Белки, жиры, углеводы и нуклеиновые кислоты - крупные полимерные молекулы, синтезируемые из мономеров в реакциях поликонденсации, протекающих с поглощением энергии. Именно поэтому для восполнения своей биомассы бактериям, помимо источника углерода, требуется источник энергии. Энергия запасается бактериальной клеткой в форме молекул аденозинтрифосфорной кислоты (АТФ). Организмы, для которых источником энергии является свет, называются фототрофами, а организмы, получающие энергию за счет окислительно-восстановительных реакций, - хемотрофами. Среди хемотрофов выделяют литотрофы (от греч. lithos - «камень»), способные использовать неорганические доноры электронов (Н2, NH3, H2S, Fe 2+ и др.), и органотрофы, использующие в качестве доноров электронов органические соединения. Бактерии, изучаемые в медицинской микробиологии, являются гетерохемоорганотрофами. Отличительная особенность этой группы - источник углерода у них является источником энергии. Среди бактерий выделяют сапрофиты (от греч. sapros - «гнилой», phyton - «растение»), питающиеся мертвым органическим материалом и независящие от других организмов, и паразиты (от греч. parasites - «нахлебник») - гетеротрофные микрорганизмы, зависимые в получении питательных веществ от макроорганизма.
Паразиты бывают факультативными, существующими как внутри макроорганизма, так и культивируемыми на искусственных питательных средах, и облигатными, которые не могут жить вне клеток макроорганизма; к ним относятся риккетсии, хламидии. 4.1.3. ФЕРМЕНТЫ БАКТЕРИЙ В основе всех метаболических реакций в бактериальной клетке лежит деятельность ферментов, являющихся биологическими катализаторами и ускоряющих химические реакции. Ферменты, образуемые бактериальной клеткой, могут локализоваться как внутри клетки - эндоферменты, так и выделяться в окружающую среду - экзоферменты. Экзоферменты играют большую роль в обеспечении бактериальной клетки доступными для проникновения внутрь источниками углерода и энергии: выделяясь в окружающую среду, они расщепляют крупные молекулы пептидов, полисахаридов, липидов до мономеров, способных проникнуть внутрь клетки. Ряд экзоферментов, например, гиалуронидаза, коллагеназа и др., являются ферментами агрессии. Ферменты также участвуют в процессах переноса веществ в бактериальную клетку. Ферментативный спектр является таксономическим признаком, характерным для семейства, рода и в некоторых случаях - для видов. Именно поэтому определением спектра ферментативной активности пользуются при установлении таксономического положения бактерий. Наличие экзоферментов можно определить с помощью дифференциальнодиагностических сред (см. раздел 4.2). 4.1.4. ПОЛУЧЕНИЕ ЭНЕРГИИ БАКТЕРИЯМИ Энергия в бактериальной клетке накапливается в форме молекул аденозинтрифосфорной кислоты, образующейся в реакциях окисления и восстановления, сопряженных с реакциями фосфорилирования. Бактерии могут получать энергию двумя способами: дыханием и брожением. Дыхание - процесс получения энергии в реакциях окисления и восстановления, сопряженных с реакциями окислительного фосфорилирования, при котором донорами электронов могут быть органические (у органотрофов) и неорганические (у литотрофов) соединения, а акцепторами - только неорганические соединения. Акцепторами электронов в процессе дыхания у бактерий могут быть кислород (в этом случае говорят о кислородном дыхании, или биологическом окислении), а также нитрат и сульфат. Конечными продуктами дыхания являются СО2 и вода. Окисляться могут углеводы, липиды, белки. Процесс окисления белков называется гниением. В процессе гниения происходит минерализация белка, при которой он разлагается до СО2, NH3, H2S. К гнилостным бактериям относятся Proteus, Pseudomonas, Bacillus cereus. Брожение, или ферментация, - процесс получения энергии, при котором как донором, так и акцептором электронов являются органические соединения. Кислород в процессе брожения участия не принимает. Восстановленные органические соединения выделяются в питательную среду и накапливаются в ней. Ферментироваться могут углеводы, аминокислоты (за исключением ароматических), пурины, пиримидины, многоатомные спирты. Не способны сбраживаться ароматические углеводороды, стероиды, каротиноиды, жирные кислоты. Эти вещества разлагаются и окисляются только в присутствии кислорода, а в анаэробных условиях они стабильны. Продукты брожения - кислоты, газы, спирты. Исходя из природы конечных продуктов, различают несколько типов брожения углеводов: спиртовое, молочнокислое, муравьинокислое, маслянокислое.
Продукты брожения используют в пищевой промышленности, а также для идентификации бактерий. 4.1.5. ОТНОШЕНИЕ К МОЛЕКУЛЯРНОМУ КИСЛОРОДУ По отношению к кислороду, а также по использованию его в процессах получения энергии микроорганизмы подразделяются на три группы: облигатные аэробы, облигатные анаэробы, факультативные анаэробы. — Облигатные аэробы растут и размножаются только в присутствии кислорода, используя его для получения энергии путем кислородного дыхания. Облигатные аэробы подразделяются на строгие, растущие при парциальном давлении атмосферы воздуха, и микроаэрофилы, использующие кислород в процессах получения энергии, но растущие только при его пониженном парциальном давлении. — Облигатные анаэробы не используют кислород для получения энергии и подразделяются на две группы: строгие и аэротолерантные. Для строгих анаэробов молекулярный кислород токсичен, он убивает микроорганизмы или ограничивает их рост. Энергию строгие анаэробы получают маслянокислым брожением. К строгим анаэробам относятся, например, некоторые клостридии (С. botulinum, С. tetani), бактероиды. Аэротолерантные анаэробы не используют кислород для получения энергии, но могут существовать в его атмосфере. К ним относятся молочнокислые бактерии. — Факультативные анаэробы растут и размножаются как в присутствии кислорода, так и без него. Энергию они получают в присутствии кислорода за счет кислородного дыхания, а при его отсутствии - брожением. У этой группы бактерий встречается также бескислородное нитратное дыхание. Различное отношение микроорганизмов к кислороду связано с наличием или отсутствием у них защитных систем, позволяющих существовать в атмосфере кислорода. В процессе окисления в присутствии кислорода образуются токсические продукты, например, перекись водорода (Н2О2). Микроорганизмы, способные существовать в атмосфере кислорода, имеют защитные механизмы, нейтрализующие токсические соединения. Например, фермент каталаза разлагает перекись водорода на воду и молекулярный кислород. Строгие анаэробы не имеют каталазы, для их культивирования создаются условия, позволяющие удалять атмосферный кислород. 4.1.6. КОНСТРУКТИВНЫЙ МЕТАБОЛИЗМ Основные органические компоненты бактериальной клетки синтезируются в реакциях полимеризации из строительных блоков: аминокислот, фосфатов, сахаров, пуриновых и пиримидиновых оснований, органических кислот. Поставщики этих строительных блоков - промежуточные продукты основных путей энергетического метаболизма.
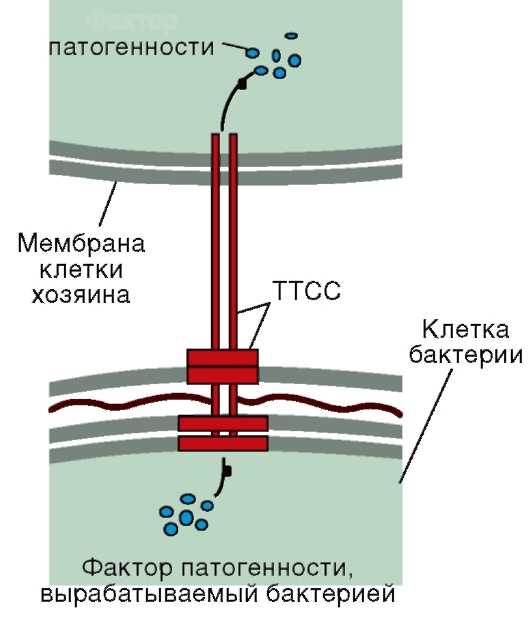
Синтез белка у бактерий происходит так же, как и в клетках эукариот и на рибосомах. Процесс синтеза белка - важная мишень для разнообразных антибактериальных препаратов. Среди бактерий выделяется группа прототрофов, способных синтезировать все компоненты клетки из одного источника углерода и энергии. Если бактерии теряют способность образовывать какоелибо жизненно важное вещество (аминокислоту, витамин), участвующее в биосинтетических процессах, то для их роста и размножения требуется его поступление в готовом виде. Такие вещества называют факторами роста, а бактерии, для роста которых они требуются, - ауксотрофами. 4.1.7. ТРАНСПОРТ ВЕЩЕСТВ Транспорт веществ в бактериальную клетку Ответственность за поступление в клетку питательных веществ лежит на ЦПМ. Существуют два типа переноса веществ в бактериальную клетку: пассивный и активный. При пассивном переносе вещество проникает в клетку только по градиенту концентрации, затраты энергии при этом не происходит. При активном переносе вещество проникает в клетку против градиента концентрации с помощью белка-переносчика - пермеазы, при этом происходит затрата энергии. Транспорт веществ из бактериальной клетки В процессе жизнедеятельности бактериям требуется выделять в окружающую среду различные белки и ферменты. Они участвуют в построении клеточных оболочек, жгутиков, пилей, расщепляют крупные полимерные молекулы, используемые в качестве питательных веществ, до размеров, способных проходить через бактериальную ЦПМ. У грамположительных бактерий белки секретируются непосредственно во внешнюю среду. У грамотрицательных бактерий из-за наличия наружной мембраны сформировались специальные секреторные системы, через которые выделение из бактериальной клетки веществ происходит с поглощением энергии. Особый интерес для медицинской микробиологии представляет III путь секреции, также используемый для направленной доставки в эукариотическую клетку бактериальных белков. В результате действия последних в клетке возникают нарушения различных функций, приводящие в конечном итоге к возникновению у человека заболеваний. Подробно III путь секреции рассмотрен в подразделе 7.2.1 (рис. 4.1). Фактор
Рис. 4.1. Схема строения III типа секреторной системы, непосредственно доставляющей в эукариотическую клетку бактериальные белки Некоторые этапы секреции могут подавляться антибактериальными препаратами. Например, транспорт через мембрану молекул, необходимых для синтеза пептидогликана, блокируется гликопептидным антибиотиком - ванкомицином. 4.1.8. РОСТ И РАЗМНОЖЕНИЕ Рост бактериальных клеток связан с синтезом и накоплением всех компонентов, входящих в ее состав, и увеличением размера в пределах, характерных для данного вида. В условиях, обеспечивающих рост микроорганизмов, происходит и процесс их деления. Для большинства бактерий характерно поперечное бинарное деление, приводящее к образованию двух дочерних клеток.
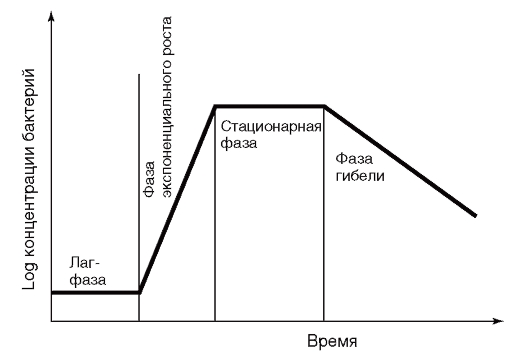
Деление бактериальной клетки начинается спустя некоторое время после завершения цикла репликации хромосомы, протекающего у бактерий по полуконсервативному механизму. Это означает, что каждая из двух нитей ДНК хромосомы служит матрицей для синтеза комплементарной дочерней цепи ДНК. Размножение бактерий бинарным делением приводит к росту числа бактериальных клеток в геометрической прогрессии. В питательной среде бактерии растут и размножаются до тех пор, пока содержание какого-нибудь из необходимых компонентов среды не достигает минимума, после чего рост и размножение прекращаются. Если на протяжении всего этого времени не прибавлять питательные вещества и не удалять конечные продукты обмена, образуется статическая бактериальная культура. Построив график, по оси абсцисс которого отложить время, а по оси ординат - количество клеток, можно получить кривую роста, описывающую зависимость числа образующихся клеток от времени размножения (рис. 4.2).
Рис. 4.2. Кривая бактериального роста На кривой роста бактерий в жидкой питательной среде можно различить несколько фаз, сменяющих друг друга в определенной последовательности. — Начальная - лаг-фаза (от англ. lag - «отставать»), охватывает промежуток времени между посевом бактерий и началом размножения. Во время лаг-фазы происходит адаптация бактериальных клеток к новым условиям культивирования. — Экспоненциальная (логарифмическая) фаза характеризуется постоянной максимальной скоростью деления клеток. Время удвоения клеток называется временем генерации, отличающимся у разных видов бактерий: у бактерий рода Pseudomonas оно составляет 14 мин, а у Mycobacterium - 24 ч. — Стационарная фаза наступает, когда количество клеток перестает увеличиваться, что связано с уменьшением содержания питательных веществ и кислорода в питательной среде и накоплением токсических продуктов обмена. Продолжительность этой фазы составляет несколько часов и зависит от вида бактерий и особенностей их культивирования. — Фаза отмирания наступает вследствие накопления кислых продуктов обмена или в результате автолиза под влиянием собственных ферментов. Продолжительность этой фазы колеблется от десятка часов до нескольких недель. На жидких питательных средах рост и размножение бактерий проявляются в виде диффузного помутнения, образования придонного осадка или поверхностной пленки. На плотных питательных средах бактерии образуют скопления клеток - колонии, считающиеся потомком одной клетки. Колонии различаются формой, размером, поверхностью, прозрачностью, консистенцией и окраской. Колонии с гладкой блестящей поверхностью называют S-формами (от англ. smooth - «гладкий»), а колонии с матовой шероховатой поверхностью - R-формами (от англ. rough - «шероховатый»). Окраска колоний определяется способностью бактерий синтезировать пигменты. Пигменты различаются по цвету, химическому составу, растворимости и обладают защитными свойствами. Вид, форма, цвет и другие особенности колоний, а также характер роста на плотных питательных средах определяются как культуральные свойства бактерий и учитываются при их идентификации. Помимо бинарного деления, некоторые представители царства Procaryotae имеют иные способы размножения. Актиномицеты могут размножаться путем фрагментации гифов. Представители семейства Streptomycetaceae размножаются спорами. Микоплазмы, помимо поперечного деления, могут размножаться почкованием. Хламидии не обладают способностью к бинарному делению. Они проходят через цикл развития, который предусматривает существование двух форм: внеклеточной - в виде инфекционных, малых размеров элементарных телец, не способных к бинарному делению, и внутриклеточной - в виде метаболически активных, крупных размеров ретикулярных телец, способных к бинарному делению. В результате бинарного деления ретикулярного тельца формируются дочерние элементарные тельца, выделяющиеся из клетки. Некоторые спирохеты, например Treponema pallidum, способны образовывать в неблагоприятных условиях цисты, которые, распадаясь на зерна, дают потомство новым бактериальным клеткам. 4.2. КУЛЬТИВИРОВАНИЕ БАКТЕРИЙ Для культивирования бактерий необходимы следующие условия. — Полноценная питательная среда. Искусственные питательные среды должны отвечать следующим требованиям: ◊ каждая питательная среда должна содержать воду, так как в ней протекают все процессы жизнедеятельности бактерий; ◊ для культивирования гетероорганотрофных бактерий в среде должны содержаться органические источники углерода и энергии (углеводы, аминокислоты, органические кислоты, липиды); ◊ для биосинтетических процессов бактериям требуются источники азота, серы, фосфаты, микроэлементы. Источником азота служит пептон - продукт неполного гидролиза белков, поставляющий аминокислоты для построения бактериальных белков. Бактерии способны утилизировать минеральные вещества в виде неорганических солей; ◊ поддержание определенного рН имеет значение для предотвращения гибели микроорганизмов от ими же образованных продуктов обмена; ◊ среда должна обладать определенным осмотическим давлением. Большинство бактерий способны расти на изотонических средах, чего достигают добавлением NaCl в концентрации 0,87%; ◊ питательные среды должны быть стерильными. В зависимости от консистенции питательные среды могут быть жидкими, полужидкими и плотными. Плотность среды достигается добавлением агара. Агар - полисахарид, получаемый из водорослей. Он плавится при температуре 100 °C, а при охлаждении остывает при 45-50 °C. Агар добавляют в концентрации 0,5% для создания полужидких сред и в концентрации 1,5-2% - для плотных. В зависимости от состава и цели применения различают простые, сложные, элективные, дифференциально-диагностические и комбинированные среды. К простым средам относятся пептонная вода, питательный бульон, мясопептонный агар. На основе этих простых сред готовят сложные среды, например, сахарный и сывороточный бульоны, кровяной агар. В зависимости от назначения среды подразделяются на элективные, среды обогащения и дифференциально-диагностические. На элективных средах лучше растет какой-то определенный микроорганизм. Например, щелочной агар, имеющий рН = 9,0, служит для выделения холерного вибриона. Другие бактерии, в частности кишечная палочка, из-за высокого рН на этой среде не растут. Среды обогащения стимулируют рост какого-то определенного микроорганизма, ингибируя рост других. Например, среда, содержащая селенит натрия, стимулирует рост бактерий рода Salmonella, ингибируя рост кишечной палочки. Дифференциально-диагностические среды служат для изучения ферментативной активности бактерий. Они состоят из простой питательной среды с добавлением субстрата, на который должен подействовать фермент, и индикатора, меняющего свой цвет в результате ферментативного превращения субстрата. Например, среды Гиса (рис. 4.3 а) используют для изучения способности бактерий ферментировать сахара; среда Клиглера позволяет дифференцировать бактерии по способности ферментировать глюкозу, лактозу и продуцировать сероводород (рис. 4.3 б). Комбинированные среды сочетают элективную среду, подавляющую рост сопутствующей микрофлоры, и дифференциальную среду, диагностирующую ферментативную активность выделяемого микроорганизма. Примерами таких сред служат среда Плоскирева и висмут-сульфитный агар, используемые при выделении патогенных кишечных бактерий. Эти среды ингибируют рост кишечной палочки. — Температура культивирования. Температура влияет на скорость размножения. По отношению к температуре роста бактерии разделяются на три основные группы: психрофилы, мезофилы и термофилы. Психрофилы живут и размножаются при пониженных температурах (от -10 до +20 °C). Мезофилы имеют температурный диапазон роста между 30 и 45 °C. К этой группе относится большинство возбудителей болезней человека, оптимальный рост которых возможен при температуре 37 °C. Термофилы способны расти при повышенных температурах. Для поддержания требуемой температуры используют специальные приборы - термостаты. — Атмосфера культивирования. Для роста и размножения строгих аэробов необходим кислород. Аэробы хорошо растут на поверхности агара на чашках Петри или в тонком верхнем слое жидкой среды. Факультативные анаэробы культивируют в аналогичных условиях.
Рис. 4.3 а. Дифференциально-диагностические среды. Среды Гиса
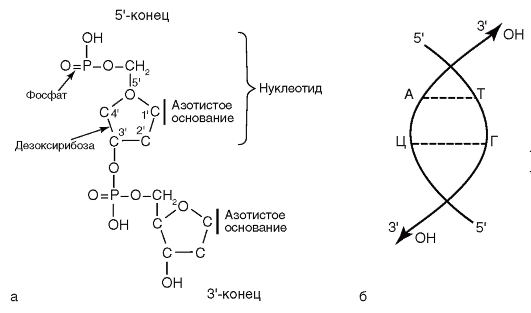
Рис. 4.3 б. Дифференциально-диагностические среды. Рост представителей семейства Enterobacteriaceae на среде Клиглера: 1 - среда до посева; 2 - Salmonella; 3 - Escherichia; 4 - Shigella; 5 - Salmonella Typhi Микроаэрофилы, нуждающиеся в пониженной концентрации кислорода, культивируют в атмосфере 5% в специальных СО2-инкубаторах или же посевы помещают в эксикаторы, в которых устанавливают горящую свечу. Облигатные анаэробы для своего роста и размножения требуют исключения доступа кислорода воздуха. Этого достигают следующими мерами: ◊ добавлением к питательным средам редуцирующих кислород веществ - тиогликолевой, аскорбиновой кислот; ◊ освобождением от кислорода воздуха жидких питательных сред путем их кипячения с последующим плотным закупориванием сосудов, в которые налиты среды, резиновыми пробками; ◊ использованием поглотителей кислорода, помещая их в герметически закрываемые емкости - «Газпаки»; этот метод используют для культивирования аэротолерантных бактерий; ◊ механическим удалением кислорода воздуха с последующим заполнением емкости инертным газом (для этих целей используют анаэростаты и анаэробные боксы). Время культивирования зависит от времени генерации. Большинство бактерий культивируют для получения видимого роста в течение 18-48 ч. Для культивирования возбудителя коклюша требуется 5 сут, а для М. tuberculosis - 3-4 нед. — Освещение. Для выращивания фототрофных микроорганизмов необходим свет. Некоторые условно патогенные микобактерии в зависимости от освещенности образуют пигмент, который используют при их идентификации. Культивирование абсолютных внутриклеточных паразитов, бактерий, относящихся к родам Rickettsia, Chlamydia, осуществляют на культурах клеток или в организме животных и членистоногих, а также на куриных эмбрионах. 4.3. СТРОЕНИЕ И РЕПЛИКАЦИЯ ГЕНОМА БАКТЕРИЙ Наследственную функцию в бактериальной клетке выполняет ДНК. Молекула ДНК построена из двух полинуклеотидных цепочек. Каждый нуклеотид состоит из азотистого основания, сахара дезоксирибозы и фосфатной группы. Азотистые основания представлены пуринами (аденином, гуанином) и пиримидинами (тимином, цитозином). Каждый нуклеотид обладает полярностью. У него имеется дезоксирибозный З'-конец и фосфатный 5'-конец. Нуклеотиды соединяются в полинуклеотидную цепочку посредством фосфодиэфирных связей между 5'-концом одного нуклеотида и 3'-концом другого (рис. 4.4). Соединение цепей обеспечивается водородными связями между комплементарными азотистыми основаниями: аденина с тимином, гуанина с цитозином. Наследственная информация у бактерий хранится в форме последовательности нуклеотидов в ДНК, которые определяют последовательность аминокислот в белке. Каждому белку соответствует определенный ген - дискретный участок ДНК, отличающийся специфичной последовательностью нуклеотидов. Совокупность всех генов называется геномом (генотипом), а внешнее проявление генома - фенотипом. Размеры генома определяются количеством нуклеотидных пар оснований.
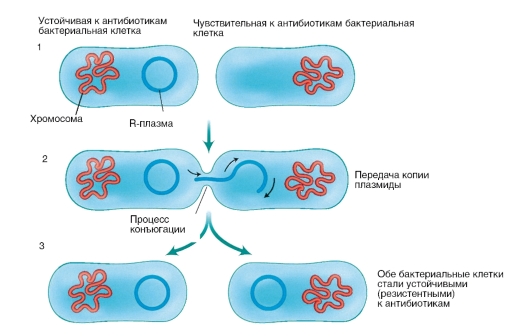
Рис. 4.4. Строение ДНК и ее элементов Бактериальный геном состоит из генетических элементов, способных к самостоятельной репликации (воспроизведению), - репликонов. Репликонами являются бактериальная хромосома и плазмиды. Геном бактерий имеет гаплоидный набор генов. Бактериальная хромосома состоит из одной двухцепочечной молекулы ДНК. Она может быть в виде одной кольцевой молекулы ДНК, как у Е. coli, или в виде линейной ДНК (B. burgdorferi). У некоторых бактерий (V. cholerae, Brucella spp.) имеются две кольцевые хромосомы. Бактериальная хромосома формирует нуклеоид бактериальной клетки и кодирует жизненно важные для бактериальной клетки функции. Плазмиды - двухцепочечные молекулы ДНК размером от 10 3 до 10 6 нуклеотидных пар, кольцевой или линейной формы. Плазмиды кодируют не основные для жизнедеятельности бактериальной клетки функции, но придающие бактерии преимущества при попадании в неблагоприятные условия существования. Некоторые плазмиды: трансмиссивные, способны передаваться из одной клетки в другую, иногда даже принадлежащую иной таксономической единице. Особое значение в медицинской микробиологии имеют плазмиды, обеспечивающие устойчивость бактерий к антибиотикам, которые получили название R-плазмид, и плазмиды, обеспечивающие продукцию факторов патогенности, способствующие развитию инфекционного процесса в макроорганизме. R-плазмиды (от англ. resistance - «противодействие») содержат гены, детерминирующие синтез ферментов, разрушающих антибактериальные препараты (например, антибиотики). В результате наличия такой плазмиды бактериальная клетка становится устойчивой (резистентной) к действию целой группы лекарственных веществ, а иногда и к нескольким препаратам. Многие R-плазмиды являются трансмиссивными: распространяясь в популяции бактерий, они делают ее недоступной к воздействию антибактериальных препаратов (рис. 4.5). Бактериальные штаммы, несущие R-плазмиды, очень часто являются этиологическими агентами внутрибольничных инфекций (см. главу 15).
Рис. 4.5. Передача R-плазмиды между бактериальными клетками Плазмиды, детерминирующие синтез факторов патогенности, в настоящее время обнаружены у многих бактерий, являющихся возбудителями инфекционных заболеваний человека. Репликация плазмид происходит независимо от хромосомы с участием того же набора ферментов, который осуществляет репликацию бактериальной хромосомы. Перед репликацией матричная двухцепочечная ДНК должна быть расплетена на отдельные цепи, что осуществляется при участии специальных ферментов, в частности ДНК-гиразы. Синтез новых цепей ДНК осуществляется ферментом ДНК-полимеразой. ДНК-полимераза способна присоединять комплементарные матрице нуклеотиды к свободному 3'-концу растущей цепи. Именно поэтому для реакции полимеризации нуклеотидов на матрице родительской цепи полимеразе требуется затравка - праймер (от англ. primer -«запал»). Праймер представляет собой короткую нуклеотидную цепочку РНК, комплементарную матричной цепи, со свободным 3'-концом. Достраивание осуществляется присоединением к свободной гидроксильной группе 3'-конца затравки нового нуклеотида. Синтез ДНК начинается с синтеза РНК. РНК-затравку для синтеза ДНК образует специальный фермент, способный инициировать синтез РНК по одноцепочечной ДНК матрицы при отсутствии затравки. После того как цепь ДНК начала синтезироваться, РНК-затравка удаляется, а образуемые бреши застраиваются ДНК-полимеразой с высокой точностью. Механизм синтеза ДНК составляет основу важного диагностического метода - полимеразной цепной реакции (ПЦР) которая подробно рассмотрена в главе 9. 4.4. ОСОБЕННОСТИ ФИЗИОЛОГИИ ГРИБОВ И ПРОСТЕЙШИХ Грибы растут в широких диапазонах температур (оптимально - при 25-30 °C), имеют половой и бесполый способы размножения, поэтому они широко распространены в окружающей среде, особенно в почве. Вместе с сине-зелеными водорослями они образуют симбиоз в виде лишайника. В этом симбиозе грибы поглощают воду и растворимые в ней вещества, а сине-зеленые водоросли поставляют грибам органические соединения. Другой вид взаимоотношений - микориза, т.е. симбиоз грибов и корней высших растений. Грибы культивируют в течение нескольких суток на сусле-агаре или жидком сусле, средах Сабуро, Чапека и др. Для этой цели можно использовать лабораторных животных. Некоторые грибы обладают диморфизмом, т.е. способностью образовывать нитчатые и дрожжевые формы в зависимости от условий роста. Дрожжеподобные формы часто образуются in vivo (при инфицировании человека грибами). Простейшие имеют органы движения (жгутики, реснички, псевдоподии), питания (пищеварительные вакуоли) и выделения (сократительные вакуоли). По типу питания они могут быть гетеротрофами или аутотрофами, а по отношению к кислороду - аэробами или факультативными анаэробами. Размножаются бесполым и половым путями. Некоторые простейшие имеют сложный жизненный цикл, сопровождаемый сменой форм развития (полового и бесполого размножения), и образуют цисты. Многие простейшие (дизентерийная амеба, лямблии, трихомонады, лейшмании, балантидии) могут расти на питательных средах, содержащих нативные белки и аминокислоты. Для их культивирования используют культуры клеток (тканей), куриные эмбрионы и лабораторных животных. 4.5. ФИЗИОЛОГИЯ ВИРУСОВ И ИХ КУЛЬТИВИРОВАНИЕ Вирусы - облигатные внутриклеточные паразиты: размножаются только в цитоплазме или ядре инфицированных клеток. Взаимодействие вируса с инфицированной клеткой может приводить к различным последствиям: — разрушению клетки (некрозу или апоптозу по типу программированной клеточной гибели), в результате чего наблюдается цитопатический эффект - клетки округляются, отделяются от соседних клеток, образуя многоядерные гигантские клетки, вакуоли и включения; — латентной инфекции, когда вирус находится внутри клетки, но не разрушает ее или трансформирует клетку организма в раковую клетку. Различают три типа взаимодействия вируса с клеткой: продуктивный, абортивный и интегративный. Продуктивный тип происходит в результате размножения вируса, т.е. его репродукции, завершающейся воспроизводством вирусного потомства - многочисленных вирионов. Продуктивный тип осуществляется в несколько стадий (рис. 4.6), таких как: — адсорбция вириона на клетке; — проникновение вириона в клетку, «раздевание» и высвобождение вирусного генома (стадия депротеинизации - в клетке вирус лишается многих своих белков); конечными продуктами депротеинизации могут быть нуклеиновая кислота, нуклеопротеид (нуклеокапсид) или сердцевина вириона; — синтез вирусных компонентов; — сборка реплицированной нуклеиновой кислоты и новых капсидных белков; — выход вирионов из клетки. В результате зараженные клетки гибнут (цитоцидное, или взрывное, действие, вызванное простыми вирусами). Сложные вирусы выходят из клеток почкованием, не разрушая их (нецитоцидное действие). Абортивный тип не завершается образованием новых вирионов, поскольку инфекционный процесс в клетке прерывается на одном из этапов.
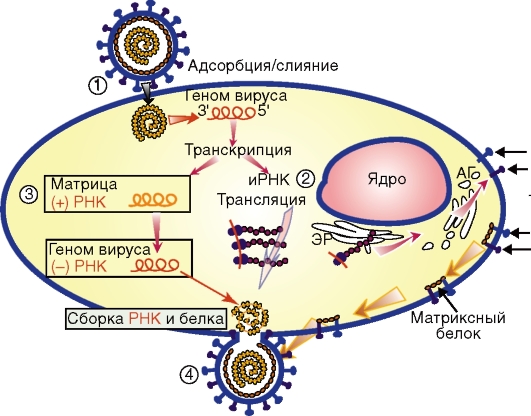
Рис. 4.6. Репродукция минус-однонитевой РНК вируса (парамиксовируса). Вирус связывается гликопротеинами своей оболочки с клеткой и сливается с плазмалеммой (1). С геномной минус-нити РНК-вируса транскрибируются: неполные плюс-нити РНК, являющиеся иРНК (2) для отдельных белков; полная минус-нить РНК - матрица для синтеза геномной минус-нити РНК вируса (3). Нуклеокапсид связывается с матриксным белком и гликопротеинмодифицированной плазмалеммой. Вирионы этого сложного вируса выходят почкованием (4) Интегративный тип, или вирогения, характеризуется встраиванием (интеграцией) вирусной ДНК в виде провируса в хромосому клетки и их сосуществованием (совместной репликацией). Провирус реплицируется в составе хромосомы и переходит в геном дочерних клеток. Под влиянием некоторых факторов провирус может исключаться из хромосомы клетки и переходить в автономное состояние с воспроизводством потомства - вирионов. Культивирование вирусов Вирусы культивируют путем заражения лабораторных животных, куриных эмбрионов и культур клеток (тканей). Присутствие вируса в исследуемом материале определяют с помощью методов индикации и идентификации. Индикация вирусов, т.е. неспецифическое обнаружение факта инфицирования, основана на выявлении биологических свойств вирусов и особенностей их взаимодействия с чувствительными клетками. Идентификация означает установление вида или типа вируса. Она осуществляется в основном с помощью реакций иммунитета или молекулярно-генетических методов (см. главу 9). Лабораторных животных (белых мышей, хомячков, кроликов, обезьян и др.) заражают вируссодержащим материалом различными способами в зависимости от тропизма вирусов (подкожно, внутримышечно, интраназально, интрацеребрально и т. д.). Наличие вирусов выявляют по развитию у животных клинических признаков заболевания, патоморфологическим изменениям органов и тканей, а также на основании реакции гемагглютинации (РГА) с вируссодержащим материалом. РГА основана на способности многих вирусов склеивать (агглютинировать) эритроциты своими гликопротеиновыми шипами (гемагглютининами). Куриные эмбрионы (5-12-дневные) заражают исследуемым материалом в различные полости и ткани зародыша. Свидетельством репродукции вирусов в куриных эмбрионах являются: — специфические поражения оболочек и тела эмбриона; — гибель эмбриона; — положительная РГА с вируссодержащей жидкостью, полученной из полостей зараженного зародыша.
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-04-05; просмотров: 141; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.142.197.198 (0.08 с.) |