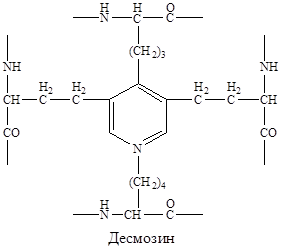Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Этапы синтеза и созревания коллагенаСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Синтез и созревание коллагена — сложный многоэтапный процесс, начинающийся в клетке, а завершающийся в межклеточном матриксе: 1. На полисомах ЭПР синтезируются полипептидные препро-α-цепи коллагена. Они содержит начиная с N-конца: 1). гидрофобный «сигнальный» пептид, содержащий около 100 АК; 2). N-концевой пропептид, содержащий около 100 АК, в том числе цистеин; 3). α-цепь коллагена 4). С-концевой пропептид, содержащий около 250 АК, в том числе цистеин. Концевые С- и N-пропептиды формируют глобулярные домены и необходимы для правильного формирования тройной спирали. «Сигнальный» пептид, обеспечивает поступление синтезируемой на рибосоме препро-α-цепи в полость ЭПР. 2. В полости ЭПР при отщеплении сигнального пептида препро-α-цепи коллагена превращаются в про-α-цепи. 3. Поступающие в полости ЭПР про-α-цепи коллагена подвергаются модификации. а). Цистеины N-пропептидов образуют внутрицепочечные дисульфидные мостики, формируя на N-конце глобулярную структуру; б). Пролины и лизины в Y-положении (гли-х-у) про-α-цепей гидроксилируются пролил-4-гидроксилазой и лизил-5-гидроксилазой в 4-гидроксипролины (Hyp) и 5-гидроксилизины (Hyl). Некоторые пролины в Х-положениях гидроксилируются в 3-гидроксипролины пролил-3-гидроксилазой. Оксигеназы (гидроксилазы), содержат Fe2+, находятся на мембране ЭПР. Для реакции необходимы а-КГ, О2 и витамин С:
Гидроксилирование пролина необходимо для стабилизации тройной спирали коллагена, ОН-группы гидроксипролина участвуют в образовании водородных связей. Гидроксилирование лизина необходимо для последующего образования ковалентных связей между молекулами коллагена при сборке коллагеновых фибрилл. в). Гидроксилизин про-α-цепей при участии гликозилтрансфераз гликозилируется галактозой или галактозилглюкозой. В молекуле коллагена сухожилий (тип I) количество углеводов равно 6, а в моллекуле коллагена капсулы хрусталика (тип TV) — 110. Роль этих углеводных групп неясна. 4). В просвете ЭПР после отделения от рибосом про-α-цепей, 3 из них с помощью С-концевых пропептидов соединяются между собой дисульфидными мостиками (цистеины С-пропептидов образуют внутри- и межцепочечные дисульфидные мостики) и скручиваются с образованием тройной спирали проколлагена. Тройная спираль проколлагена стабилизируется водородными связями. После этого гидроксилирование и гликозилирование про-α-цепей прекращается. 5). Из ЭПР молекулы проколлагена перемещаются в аппарат Гольджи, включаются в секреторные пузырьки и секретируются в межклеточное пространство. 6). В межклеточном матриксе от некоторых проколлагенов (I, II, III, V, XI типов) проколлагенпептидазы отщепляют концевые С- и N-пропептиды, в результате чего образуется тропоколлагены. У проколлагенов IV, VIII, X типов концевые пропептиды не отщепляются. Химический состав межклеточного матрикса В состав межклеточного матрикса входят: 1). Коллагеновые и эластиновые волокна. Они придают ткани механическую прочность, препятствуя ее растяжению; 2). аморфное вещество в виде ГАГ и протеогликанов. Оно удерживает воду и минеральные вещества, препятствует сдавливанию ткани; 3). неколлагеновые структурные белки - фибронектин, ламинин, тенасцин, остеонектин и др. Кроме того, в межклеточном матриксе может присутствовать минеральный компонент - в костях и зубах: гидроксиапатит, фосфаты кальция, магния и т.д. Он придает механическую прочность костям, зубам, создает запас в организме кальция, магния, натрия, фосфора. Функция межклеточного матрикса Межклеточный матрикс выполняет в организме разнообразные функции: · образует каркас органов и тканей; · является универсальным «биологическим» клеем; · участвует в регуляции водно-солевого обмена; · образует высокоспециализированные структуры (кости, зубы, хрящи, сухожилия, базальные мембраны). · окружая клетки, влияет на их прикрепление, развитие, пролиферацию, организацию и метаболизм.
Эластин — основной белок эластических волокон, которые в больших количествах содержатся в межклеточном веществе кожи, стенок кровеносных сосудов, связках, лёгких. Эти ткани могут растягиваться в несколько раз по сравнению с исходной длиной, сохраняя при этом высокую прочность на разрыв. Строение эластина Эластин — гликопротеин с молекулярной массой 70 кДа. Первичная структура эластина образована полипептидной цепью из 800 АК, в которой преобладают глицин, валин, аланин, содержится много пролина и лизина, немного гидроксипролина, отсутствует гидроксилизин. Большое количество гидрофобных радикалов препятствует созданию регулярной вторичной и третичной структуры эластина, поэтому он приобретает различные конформации. В межклеточном пространстве молекулы эластина образуют волокна и слои, в которых отдельные пептидные цепи связаны множеством жёстких поперечных сшивок в разветвлённую сеть. Сшивки между остатками лизина двух, трёх или четырёх пептидных цепей, образуют специфические структуры, которые называются десмозинами (десмозин или изодесмозин).
Кроме десмозинов, в образовании поперечных сшивок может участвовать лизиннорлейцин, который образуется двумя остатками лизина. Наличие ковалентных сшивок между пептидными цепочками с неупорядоченной, случайной конформацией позволяет всей сети волокон эластина растягиваться и сжиматься в разных направлениях, придавая соответствующим тканям свойство эластичности. Синтез эластина Эластин синтезируется фибробластами в виде растворимого мономера - «тропоэластина». В межклеточном пространстве после образования поперечных сшивок эластин приобретает свою конечную внеклеточную форму, которая характеризуется нерастворимостью, высокой стабильностью и очень низкой скоростью обмена. Нарушения структуры эластина и их последствия Снижение активности лизилоксидазы, вызванное дефицитом меди, пиридоксина или дефицит лизилоксидазы, связанный с генетическим дефектом, приводит к снижению или прекращению образования десмозинов. В результате поперечных сшивок нет или их недостаточное количество. При этом, у эластических тканей снижается предел прочности на разрыв, появляются такие нарушения, как истончённость, вялость и растяжимость. Клинически эти нарушения могут проявляться кардиоваскулярными изменениями (аневризмы и разрывы аорты, дефекты клапанов сердца), частыми пневмониями и эмфиземой лёгких. Катаболизм эластина Катаболизм эластина происходит при участии эластазы нейтрофилов. Это очень активная протеаза, которая выделяется во внеклеточное пространство нейтрофилами и разрушает эластин и другие структурные белки. Особое значение это имеет в лёгких, поскольку лёгочная ткань не регенерирует. Разрушение эластина в альвеолярных стенках ведёт к потере эластичных свойств, разрушению альвеол и развитию эмфиземы лёгких. В норме эластазу нейтрофилов и другие протеазы ингибирует α1-антитрипсин. Основное количество α1-антитрипсина синтезируется печенью и находится в крови. В лёгких α1-антитрипсин синтезируется альвеолярными макрофагами, что и обеспечивает защиту альвеол от действия эластазы. При дефиците α1-антитрипсина, который может быть следствием различных мутаций, повышается риск развития эмфиземы лёгких.
|
||||||
|
|
Последнее изменение этой страницы: 2016-08-10; просмотров: 996; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.147.75.217 (0.007 с.) |