Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Постсинтетическая модификация белкаСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Во многих случаях белки синтезируются в виде предшественников – биологически неактивных молекул.Их функциональная активность проявляется в результате превращений, называемых постсинтетической или посттрансляционной модификацией (процессинг). Примеры посттрансляционной модификации белков: - протеолитическое отщепление N-концевого формилметионина или метионина; - отщепление сигнальных пептидов; - частичный протеолиз; - посттрансляционная модификация белков по аминокислотным радикалам: ковалентное присоединение простетической группы, метилирование радикалов лизина и аргинина, ацетилирование N-концевой аминокислоты, фосфорилирование гистонов и негистоновых белков хроматина, гидроксилирование радикала пролина; присоединение олигосахаридных фрагментов (гликозилирование) к радикалам аспарагина, серина и треонина и т.д. Выбор правильной структуры белка происходи при участии белков шаперонов. Гидрофобные участки на поверхности глобулы шаперонов-70 взаимодействуют с гидрофобными участками синтезированной цепи, защищая ее от неравильных взаимодействий с другими белками цитозоля. Шапероны-60 участвуют в исправлении пространственной структуры неправильно свернутой или поврежденной цепи. Мутации в шапероне, входящем в состав хрусталика глаза, приводят к помутнению хрусталика из-за агрегации белков и развитию катаракты. Транспорт синтезированных белков через мембраны Синтезированный белок поступает из рибосомы в цитозоль. Если он не используется для нужд самой клетки, т.е. относится к экспортируемым (секретируемым) белкам, то он переносится через мембрану при помощи низкомолекулярных пептидов (15-30 аминокислотных остатков), содержащих гидрофобные радикалы. Это сигнальные пептиды. В мембране формируется канал, через который сигнальный пептид проникает внутрь цистерны эндоплазматического ретикулума, и протаскивает за собой синтезируемую молекулу белка. Под действием сигнальной пептидазы N-концевая сигнальная последовательность отщепляется, а белок через аппарат Гольджи выходит из клетки в форме секреторного пузырька.
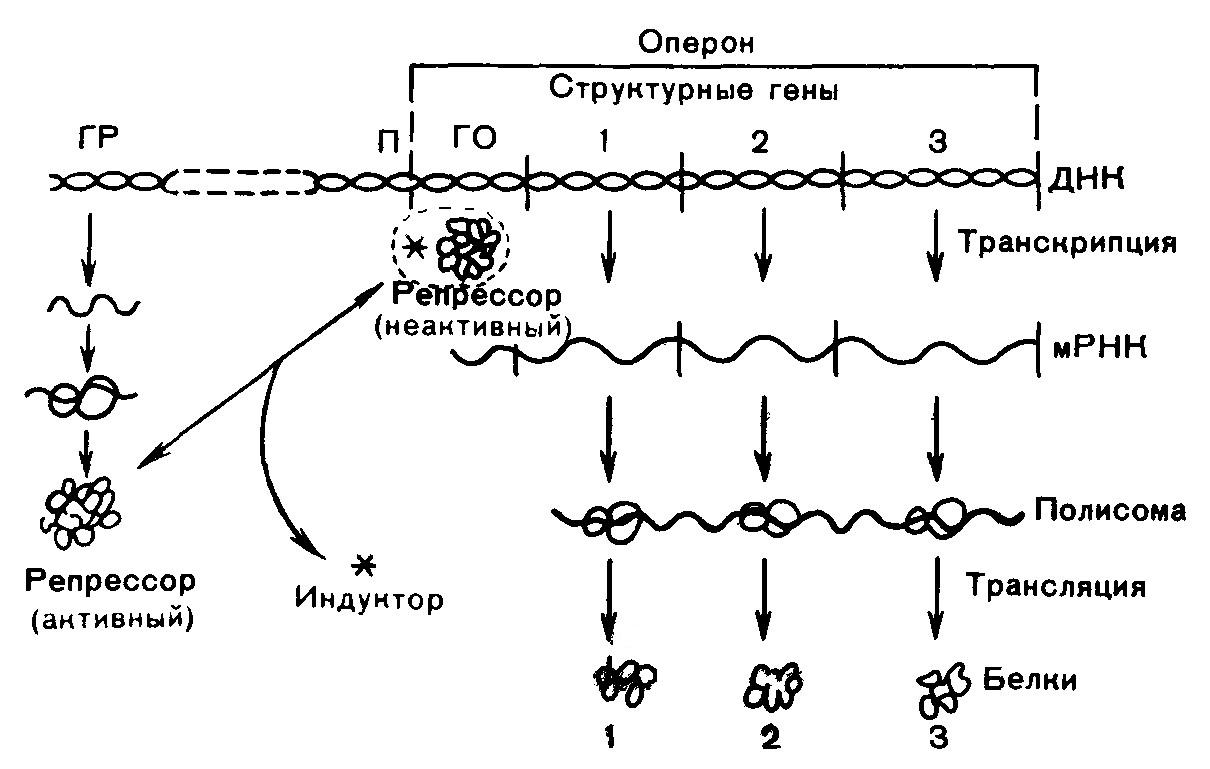
РЕГУЛЯЦИЯ СИНТЕЗА БЕЛКА Концентрация многих белков в клетке непостоянна и изменяется в зависимости от состояния клетки и внешних условий. Это происходит в результате регуляции скоростей синтеза и распада белков. В клетках млекопитающих существуют два вида регуляции биосинтеза белков: - кратковременная, обеспечивающая адаптацию организма к изменениям окружающей среды; - длительная, стабильная, определяющая дифференцировку клеток и разный белковый состав органов и тканей. Наиболее распространенным механизм регуляции синтеза белков является регуляция на уровне транскрипции (образования первичного транскрипта). Синтез в базальном состоянии называют конститутивным синтезом. Различают две формы регуляции – индукция синтеза (положительная регуляция) и репрессия синтеза (отрицательная регуляция). Понятия индукции и репрессии предполагают изменение скорости синтеза белка по отношению к исходному (базальному) уровню. Если скорость конститутивного синтеза белка высока, то белок регулируется по механизму репрессии синтеза, и, наоборот – при низкой базальной скорости наблюдается индукция синтеза. Согласно теории Ф. Жакоба и Ж. Моне, в генетическом аппарате бактериальной клетки существуют опероны – отрезки ДНК, содержащие структурные гены определенных белков (цистроны), и регуляторные участки. Считывание генетического кода начинается с промотора, расположенного рядом с геном-оператором. Ген-оператор расположен на крайнем отрезке структурного гена. Он либо запрещает, либо разрешает репликацию мРНК на ДНК. Деятельность оперона контролирует ген-регулятор. Белок-репрессор осуществляет связь опероном и геном-регулятором. Репрессор образуется в рибосомах ядра на мРНК, синтезированной на гене-регуляторе. Он образует комплекс с геном-оператором и блокирует синтез мРНК, а, следовательно, и белка. Репрессор может связываться с низкомолекулярными веществами – индукторами, или эффекторами. После этого он теряет способность связываться с геном-оператором, ген-оператор выходит из-под контроля гена-регулятора, и начинается синтез мРНК. Это индукция синтеза (рис. 14).
У эукариот механизмы регуляции синтеза белка более сложные. Преобладают положительные регуляторные механизмы. Основной регуляторной точкой является стадия инициации транскрипции. Регуляторные элементы, стимулирующие транскрипцию, называют энхансерами, а подавляющие ее – сайленсерами. Они могут располагаться как рядом с промотором, так и на удалении от него, и избирательно соединяться с белками-регуляторами: энхансеры – с белками-индукторами, сайленсеры – с белками-репрессорами. Процесс взаимодействия регуляторных элементов с белками-регуляторами регулируется сигнальными молекулами – гормонами, некоторыми метаболитами. Знание особенностей структуры и функционирования рибосом прокариот и эукариот позволило разработать новые типы антибиотиков. Синтез нуклеиновых кислот и белков является ключевым процессом, необходимым для поддержания жизнедеятельности клетки. Если его каким-либо образом выключить - клетка погибнет. Существуют лекарственные препараты, нарушающие синтез пуриновых оснований и аминокислот, синтез нуклеиновых кислот (рис. 16), синтез белка на различных уровнях только в клетках бактерий (таблица 4).
Рис 16. Препараты, нарушающие синтез нуклеиновых кислот
Таблица 4 Средства, угнетающие синтез белков бактериальной клетки
ГЕННАЯ ИНЖЕНЕРИЯ Генная инженерия - система экспериментальных приемов, позволяющих конструировать лабораторным путем (в пробирке) искусственные генетические структуры в виде так называемых рекомбинантных или гибридных молекул ДНК. Технология рекомбинантных ДНК использует следующие методы: 1. Специфическое расщепление ДНК рестриктазами. В качестве мишеней рестриктаз часто выступают палиндромы из 4-6 пар оснований - сайты рестрикции. Одни рестриктазы вносят разрывы по оси симметрии, образуются так называемые "тупые" концы. Другие - со сдвигом, образуются "липкие" концы, то есть фрагменты имеют на своих концах однонитевые взаимно комплементарные участки длиной в четыре нуклеотида. Такие фрагменты особенно удобны для создания рекомбинантных ДНК. 2. Секвенирование нуклеотидов. С помощью электрофореза фрагменты ДНК, различающиеся по размеру, можно разделить, а затем исследовать каждый фрагмент отдельно. Это позволяет построить рестрикционную карту, на которой указано положение каждого сайта рестрикции относительно других участков. 3. Конструирование рекомбинантной ДНК: Для получения рекомбинантной ДНК плазмиды выделяют из Е. coli и удаляют из них часть кольцевой молекулы ДНК с помощью рестриктазы. Комплементарные цепи молекулы ДНК разрезаются в разных местах, в результате чего образуются «липкие» концы. На фрагменте ДНК, выбранном для пересадки, создают «липкие» концы, используя ту же рестриктазу. Если смешать фрагмент ДНК (ген) и плазмиду, то они соединятся «липкими» концами. Затем с помощью лигазы вновь получают кольцевую молекулу ДНК, но теперь она вместе с плазмидной ДНК содержит ген, выбранный для пересадки. Это и есть рекомбинантная ДНК. Возможны: - сшивка по одноименным "липким" концам (рестриктазно лигазный метод). Комплементарные друг другу участки имеют тенденцию к ассоциации за счет спаривания оснований. Для восстановления разрывов используют фермент ДНК-лигазу. - сшивка по "тупым" концам (коннекторный метод). Тупые концы могут быть соединены ДНК-лигазой. Эффективность реакции ниже, чем при сшивке по липким концам. - сшивка фрагментов с разноименными липкими концами. Применяют линкеры - химически синтезированные олигонуклеотиды, представляющие собой сайты рестрикции или их комбинацию. Существуют линкеры "тупой конец - липкий конец". Описанная выше процедура сложна и позволяет получать лишь очень небольшие количества рекомбинантной ДНК. 4. Клонирование (размножение) рекомбинантной ДНК: - клонирование ДНК in vivo. Если к культуре Е. coli добавить рекомбинантные плазмиды, то они могут включаться в бактериальные клетки - получаются рекомбинантные бактерии. Плазмиды в клетке начинают реплицироваться. При размножении бактерий вновь образующиеся бактериальные клетки тоже содержат эти плазмиды. Из рекомбинантных бактерий можно выделить клонированные рекомбинантные плазмиды, а из них - исследуемый фрагмент ДНК. Таким путем можно выделить ген или любой другой фрагмент ДНК в количествах, достаточных для исследовательских целей. - аплификация (увеличение числа копий) ДНК in vitro. В 1985 году К. Мюллис с сотрудниками разработали метод клонирования последовательностей ДНК in vitro, который получил название полимеразной цепной реакции (ПЦР). К анализируемому образцу ДНК добавляют в избытке 2 синтетических праймера. Праймеры ориентированы таким образом, что синтез с помощью полимеразы протекает только между ними, удваивая количество копий этого участка ДНК. Амплифицированный участок называют ампликоном. Амплификация заключается в повторяющихся циклах, представляющих собой трехступенчатый процесс: I - денатурация ДНК при 95 °С; II - отжиг праймеров с комплементарными последовательностями (40-60 °С); III - последующая достройка полинуклеотидных цепей от праймеров с помощью ДНК-полимеразы при температуре 70-75 °С (рис. 17).
Продолжительность одного цикла менее 3 мин. Таким образом, за 2 ч можно получить около миллиарда копий определяемой последовательности ДНК. Размноженный in vitro фрагмент получают в количествах, достаточных для его прямого секвенирования. ПЦР называют бесклеточным молекулярным клонированием (cell-free molecular cloning). С использованием ПЦР разработаны новые диагностические тесты на генетические и инфекционные заболевания. Метод используют для ранней диагностики наличия в организме вируса иммунодефицита человека (ВИЧ). Метод анализа индивидуальных сперматозоидов нашел практическое применение в судебной медицине. С помощью ПЦР удалось амплифицировать и клонировать фрагменты митохондриальной ДНК из ископаемых останков мозга человека возраста 7 тысяч лет. 5. Введение гена в клетку. Ген нужно ввести в клетку таким образом, чтобы он не был разрушен клеточными нуклеазами, а интегрировался с геномом клетки. Используют 2 способа. Трансдукция. Вектор - молекула ДНК или РНК, состоящая из двух компонентов: векторной части (носителя) и клонируемого чужеродного гена. Задача вектора – донести выбранную ДНК в клетку-рецепиент и встроить ее в геном. В состав вектора должен входить маркерный ген, позволяющий селектировать измененные клетки. Можно выделить 2 группы маркерных генов: - селективные гены, отвечающие за устойчивость к антибиотикам или гербицидам; - репортерные гены, кодирующие нейтральные для клеток белки, наличие которых в тканях может быть легко тестировано. За способность гена к экспрессии отвечают регуляторные последовательности, которые также необходимо встроить в векторную молекулу. Типы векторов: - бактериальные плазмиды. Одна их наиболее часто употребляемых плазмид для клонирования pBR 322 создана на основе плазмид, выделенных из E. coli; - вирусы. Есть вирусы, которые не ведут к гибели клетки, но встраиваются в геном клетки-хозяина и размножаются вместе с ней, либо вызывают ее неконтролируемый рост, т.е. превращают в раковую; - гибридные векторы, содержащие ДНК фага и плазмиды - космиды и фазмиды. 2. Прямое введение гена в клетку – трансформация. Ее виды: Трансфекция. ДНК адсорбируется на кристаллах фосфата кальция. Они поглощаются клеткой путем фагоцитоза. Микроинъекция ДНК с помощью микропипеток диаметром 0,1-0,5 микрона и микроманипулятора. Электропорация основана на том, что импульсы высокого напряжения обратимо увеличивают проницаемость биомембран. «Мини-клетки» получают путем блокирования донорных клеток в митозе колцемидом. Вокруг каждой хромосомы формируется новая ядерная мембрана. Затем их обрабатывают цитохалазином В и центрифугируют. Образуются мини-клетки - микроядра, инкапсулированные в цитоплазматическую мембрану. Для их слияния подбирают специальные мягкие условия. Упаковка в липосомы используется для защиты экзогенного генетического материала от разрушающего действия рестриктаз. Метод биологической баллистики является одним из самых эффективных на сегодняшний день методов трансформации растений. На мельчайшие частички вольфрама напыляется ДНК вектора. Они помещаются внутрь биолистической пушки, откуда с огромной скоростью выбрасываются и, разрывая клеточные стенки, входят в цитоплазму и ядро клеток. С развитием генной инженерии появилась возможность заставить микроорганизмы синтезировать вещества, которые получить другими методами сложно - интерферон, инсулин, соматостатин, соматотропин, фермент урокиназу, некоторые факторы свертывания крови и др. Все эти белки применяются для лечения болезней. Плазмиды можно ввести и в клетки эукариот. Генетическая транформация соматических клеток млекопитающих позволяет изучать тонкие механизмы регуляции экспрессии генов и целенаправленно модифицировать генетический аппарат клетки, что имеет огромное значение для медицинской генетики.
|
|||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-07-14; просмотров: 913; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.217.89 (0.014 с.) |