Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Превращение активного ацетата в мевалоновую кислоту.Содержание книги
Поиск на нашем сайте
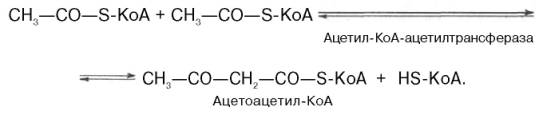
В результате конденсации 2 молекул ацетил-КоА образуется ацетоацетил-КоА, который при взаимодействии с третьей молекулой ацетил-КоА образует β-гидрокси-β-метилглутарил-КоА. Затем в ГМГ-КоА-редуктазной реакции при участии фермента ГМГ-КоА-редуктазы он восстанавливается до мевалоновой кислоты. Эта необратимая реакция лимитирует скорость биосинтеза холестерина.
Образование сквалена из мевалоновой кислоты. Мевалоновая кислота фосфорилируется при помощи АТФ и участвует в ряде реакций, в результате чего образуется фарнезилпирофосфат (С15). Затем 2 молекулы фарнезилпирофосфата конденсируются и восстанавливаются при помощи НАДФН + Н+, образуя сквален (С30):
Циклизация сквалена в холестерин. Под влиянием сквален-оксидоциклазы первоначально образуется ланостерин, который затем превращается в холестерин:
Регуляция синтеза холестерина Основным пунктом регуляции является реакция образования мевалоновой кислоты. Регуляторным ферментом является ГМГ-КоА-редуктаза. 1. Аллостерическая регуляция. Холестерин, а в печени - и желчные кислоты ингибируют ГМГ-КоА-редуктазу. 2. Репрессия синтеза ГМГ-КоА-редуктазы холестерином. 3. Регуляция путем фосфорилирования-дефосфорилирования ГМГ-КоА-редуктазы; активна нефосфорилированная форма. Глюкагон вызывает дезактивацию, а инсулин – активацию путем сложного каскада реакций. Таким образом, скорость синтеза холестерина изменяется при смене абсорбтивного и постабсорбтивного состояний. 4. Скорость синтеза ГМГ-КоА-редуктазы в печени подвержена четким суточным колебаниям: максимальна в полночь, а минимальна - в утренние часы. Обмен эфиров холестерина В клетках этерификация холестерина происходит при действии ацил-КоА-холестерин-ацилтрансферазы (АХАТ): Ацил-КоА + Холестерин ® НS-КоА + Ацилхолестерин Образуется в основном линолеилхолестерин. Эфиры находятся главным образом в цитозоле в составе липидных капель. Образование эфиров можно рассматривать, с одной стороны, как механизм удаления из мембран избыточного холестерина, а с другой - как механизм запасания холестерина в клетке. Мобилизация запасов происходит при участии ферментов эстераз, гидролизующих эфиры холестерина: Ацилхолестерин + Н2О ® Жирная кислота + Холестерин Синтез и гидролиз эфиров происходят особенно активно в клетках коры надпочечников. В липопротеинах крови образование эфиров протекает при участии лецитин-холестерин-ацилтрансферазы (ЛХАТ), катализирующей перенос ацильного остатка с лецитина на холестерин. ЛХАТ образуется в печени, секретируется в кровь и присоединяется к липопротеинам. Наиболее активна ЛХАТ в ЛВП, где она локализована в поверхностном слое. Активатором ЛХАТ является аполипопротеин А12 (АпоА1) – белок ЛВП, синтезирующийся в печени и кишечнике. Образующиеся здесь эфиры холестерина гидрофобны и погружаются в липидное ядро. В фосфолипидном монослое освобождается место для холестерина, которое может быть заполнено холестерином из клеточных мембран или из других липопротеинов. Таким образом, ЛВП в результате действия ЛХАТ оказываются ловушкой холестерина. Синтез желчных кислот В печени часть холестерина превращается в желчные кислоты. Желчные кислоты можно рассматривать как производные холановой кислоты, хотя холановая кислота как таковая в организме не образуется. Первый этап синтеза желчных кислот – гидроксилирование холестерина под действием 7α-гидроксилазы. Данная реакция является лимитирующей. В последующих реакций в гепатоцитах синтезируются первичные желчные кислоты - хенодезоксихолевая и холевая (рис. 6).
Рис. 6. Схема синтеза первичных желчных кислот После выделения желчи в кишечник при действии ферментов кишечной флоры из них образуются вторичные желчные кислоты - литохолевая и дезоксихолевая. Они всасываются в кишечнике, с кровью через воротную вену попадают в печень, а затем - в желчь. В желчи содержатся главным образом конъюгированные желчные кислоты, т. е. их соединения с глицином или таурином. Концентрация желчных кислот в желчи около 1%. Основная часть желчных кислот участвует в гепатоэнтеральной циркуляции. Небольшая часть желчных кислот - около 0,5 г за сутки - выводится с калом. Это компенсируется синтезом в печени новых желчных кислот; фонд желчных кислот обновляется примерно за 10 дней. Холестерин организма человека по происхождению подразделяют на эндогенный холестерин, синтезированный в печени и других органах т тканях, и экзогенный холестерин (холестерин пищи). Удаление холестерина из тканей происходит путем его окисления в желчные кислоты в печени с последующей их экскрецией с калом (примерно 0,5 г в сутки) и путем экскреции неизмененного холестерина (тоже с калом). В стационарном состоянии: (Холестеринэнд + Холестеринэкз) - (Холестеринэкскр + Желчные кислотыэкскр) = 0 Если представленный баланс нарушен, то концентрация холестерина в тканях и в крови изменяется. Повышение концентрации холестерина в крови – гиперхолестеринемия. При этом увеличивается вероятность заболевания атеросклерозом и желчнокаменной болезнью. РЕГУЛЯЦИЯ ЛИПИДНОГО ОБМЕНА Обмен липидов регулируется ЦНС. Длительный отрицательный эмоциональный стресс, увеличение выброса катехоламинов в кровяное русло может вызвать заметное похудение. Адреналин и норадреналин увеличивают активность тканевой липазы и скорость липолиза в жировой ткани; в результате повышается содержание жирных кислот в плазме крови. Действие глюкагона на липолитическую систему сходно с действием катехоламинов. Инсулин оказывает противоположное адреналину и глюкагону действие на липолиз и мобилизацию жирных кислот. Соматотропный гормон стимулирует липолиз, индуцируя синтез аценилатциклазы. Гипофункция гипофиза приводит к отложению жира в организме (гипофизарное ожирение). Тироксин, половые гормоны, также оказывают влияние на липидный обмен. Удаление половых желез вызывает у животных избыточное отложение жира. НАРУШЕНИЯ ЛИПИДНОГО ОБМЕНА
|
|||||||
|
|
Последнее изменение этой страницы: 2016-07-14; просмотров: 423; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.191.62.68 (0.009 с.) |