Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Особенности развития круглоротыхСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Многие черты развития круглоротых являются переходными между типом развития ланцетника и вышестоящих позвоночных, например осетровых рыб или амфибий. Так, в яйцеклетках миног желток распределен неравномерно, с преобладанием в вегетативном полушарии, а наблюдается полное неравномерное дробление, сходное с таковым амфибий. Как и у амфибий, стенки бластулы миног многослойные, а вегетативное дно сложено из большего числа клеточных слоев, чем анимальная крыша, следовательно, бластула миног является как бы зеновского узелка у птиц, что и демонстрирует глубокую гомологию между развитием птиц и рептилий. С другой стороны, наличие хорошо выраженного бластопора и присутствие у большинства рептилий (ящерицы, змеи и черепахи) округлой первичной пластинки взамен вытянутой первичной полости птиц сближают зародышей рептилий с анамниями, особенно с зародышами акуловых рыб. Развитие птиц. Классическим объектом эмбриологических исследований является куриный эмбрион. Поэтому развитие представителей класса птиц удобно рассмотреть на примере эмбриогенеза кур. Яйцеклетка у птиц богата желтком — полилецитальная. Диаметр яйцеклетки курицы достигает 3,5 см, а у страуса — до 10 см. В отложенном птичьем яйце находится не яйцеклетка, а многоклеточный зародыш на стадии бластулы или ранней гаструлы. К яйцеклетке относится только желток. Яйцеклетка, продвигаясь по яйцеводу, оплодотворяется. Диаметр овулировавшегося яйца составляет 50 мкм и постепенно увеличивается до 6 мм, а затем и до 35 мм. Желточный материал синтезируется в печени, переносится в фолликулярные клетки, а затем в цитоплазму яйцеклетки. У рептилий и птиц наблюдается полиспермия, и, хотя с женским пронуклеусом взаимодействует один мужской, избыточные спермии — мероциты (блуждающие ядра) способствуют переработке желтка. Дробление зиготы птиц неполное, неравномерное, частичное, дискоидальное, так как дробится всего 1/500 часть зиготы на анимальном полюсе. Первые три борозды дробления — радиальные, затем появляются широтные и тангенциальные полосы. При дроблении возникают бластомеры различной величины, и в конечном итоге формируется многослойная дискобластула, распластанная на нераздробившемся желтке. Периферическая зона дискобластулы называется перибластом, а центральная — зародышевым диском (бластодиском). При увеличении числа клеток в бластодиске до 100 бластомеров часть желтка под бластодиском разжижается и образуется подзародышевая полость. Морфологически, если рассматривать бластодиск сверху, можно заметить, что его периферическая часть более темная, чем центральная. Это объясняется большей толщиной периферии бластодиска. Таким образом, различают светлую центральную часть (area pellucida) и темную периферическую часть (area ораса) бластодиска. Как отмечалось выше, к моменту откладки яйца зародыш находится в первой фазе гаструляции. Морфологически это проявляется обособлением внезародышевой энтодермы путем деламинации зародышевого диска и возникновением двухслойного зародыша. Так в зародышевом диске возникают две части: эпибласт — наружная и гипобласт — внутренняя. Периферическая часть гипобласта в дальнейшем образует внезародышевую энтодерму, которая обрастает желтком, формируя желточный мешок — провизорный орган, выполняющий трофическую и кроветворную функции. В стенке желточного мешка обнаруживаются первичные половые клетки (внегонадное происхождение). Наиболее важные морфогенетические процессы развертываются в эпибласте (в пределах центральной части area pellucida), который в последние годы считается источником развития всех трех зародышевых листков. Здесь возникают клеточные потоки вследствие асинхронного дробления клеток и их направленного перемещения. Клетки перемещаются в течение 3-4 ч инкубации. Схема движения клеточных потоков выглядит следующим образом: клетки латеральных частей эпибласта (зародышевого щитка) перемещаются в хвостовую часть зародыша и, встречаясь здесь, продвигаются далее по средней линии эпибласта к головному концу. К 12 ч инкубации клеточный поток принимает вид первичной полоски. Второй поток клеток (более медленный) распространяется со стороны головного конца зародыша и направляется по средней части эпибласта навстречу первому потоку. Место встречи двух потоков обозначается как гензеновсшй, или первичный, узелок — важнейший морфологический маркер развивающегося зародыша. Передняя часть гензеновского узелка рассматривается как гомолог дорсальной губы бластопора у выше изученных представителей животных. В свою очередь, первичная полоска рассматривается гомологом латеральных губ бластопора. На вершине гензеновского узелка появляется углубление — первичная ямка, а по средней линии первичной полоски — первичная бороздка, как продолжение первичной ямки. Обе эти структуры рассматриваются как гомолог бластопора. К 16 ч инкубации все описанные структуры хорошо выявляются на фиксированных препаратах. Таким образом, в процессе развития птиц появляются структуры — морфогенетические маркеры, гомологичные таковым в эмбриогенезе низших позвоночных, доказывающие сходство эмбрионального развития разных представителей животного мира. Развитие млекопитающих В развитии низших млекопитающих (однопроходных и сумчатых) можно проследить различные стадии перехода от полилецитальных, дискоидальных дробящихся яиц рептилий к алецитальным, голобластическим яйцам высших (плацентарных) млекопитающих. У однопроходных (ехидна, утконос) яйца богаты желтком, и дробление у них дискоидальное. Дальнейшее развитие изучено недостаточно, но, по-видимому, оно более всего похоже на таковое рептилий: на бластодиске формируется клеточное скопление — первичная пластинка, а в ней — гастральный мешочек, сходный с мезодермальным мешочком рептилий и также смыкающийся с ранее отделившейся энтодермой. Дальнейшие перемещения клеток — иммиграция, характеризуют вторую фазу гаструляции. Как было отмечено ранее, первые миграционные явления наблюдаются в области, гомологичной дорсальной губе бластопора (в области переднего края гензеновского узелка). Эти клетки, подворачиваясь, проходят через ямку и распространяются в направлении головного конца зародыша, формируя головной отросток, — хорду (нотохорд). В свою очередь, хорда индуцирует раннюю детерминацию клеток нервной пластинки в составе эпибласта. Одновременно с материалом хорды из противоположного края гензеновского узелка и передней части первичной полоски в иммиграцию вовлекаются новые клетки, однако последние включаются в состав гипобласта головной части зародыша. При этом происходит деформация гензеновского узелка и укорочение первичной полоски. Все эти процессы протекают в течение 8—10 ч инкубации. Клетки оставшейся части первичной полоски представляют собой материал сомато- и спланхномезодермы. Через 15 ч инкубации эти клетки приходят в движение и мигрируют внутрь вдоль первичной полоски, распространяясь над гипобластом по бокам от удлиняющейся хорды. Таким образом, в результате иммиграции из эпибласта выселяются материал хорды, прехордальной пластинки (а, по некоторым данным, материал зародышевой энтодермы), мезодермы, что сопровождается деформацией, укорочением и исчезновением гензеновского узелка и первичной полоски. Оставшийся материал эпибласта представляет собой эктодерму, который в своем составе содержит клетки будущей нервной трубки. Как указывалось выше, хорда индуцирует развитие нервного желобка, сворачивающегося затем в трубку и погружающегося под эктодерму. Клеточный материал, который является источником развития хорды, располагается в области передней части гензеновского узелка (у низших позвоночных — в области дорсальной губы бластопора) и благодаря свойству индуцировать развитие нервной системы назван организатором. Открытие организатора первоначально у амфибий положило начало бурному расцвету экспериментальной эмбриологии, а автор открытия Г. Шпеман в 1935 г. был удостоен Нобелевской премии. Клеточный материал мезодермы дифференцируется на сомиты, латерально расположенные нефротомы и наиболее латерально лежащие спланхнотомы. Сомиты и нефротомы — это сегментированные части мезодермы, спланхнотомы — несегментированные. Последние в дальнейшем расслаиваются на висцеральный и париетальный листки со щелевидной целомической полостью между ними. Под хордой располагается энтодерма, которая сворачивается в кишечную трубку, сообщающуюся с желтком. Так возникает комплекс эмбриональных зачатков тканей. Наиболее сложна топографическая дифференцировка мезодермы. В составе сомита различают следующие части: склеротом — вентромедиальную часть, дерматом — дорсолатеральную часть и миотом — дорсомедиальную часть. Клетки склеротома интенсивно размножаются, выселяются из состава сомита, окружают хорду и дифференцируются в хрящевые клетки (возникают сложные формообразовательные процессы закладки позвонков, ребер, лопаток и других костей скелета). Дерматом является источником развития дермы кожи, а миотом — скелетной мышечной ткани. Образование внезародышевых оболочек. Параллельно образованию тела зародыша происходит образование внезародышевых оболочек, или провизорных органов. Это является важным эволюционным приспособлением для независимого существования на суше и сохранения вокруг зародыша влажной среды. Впервые яйца на суше стали откладывать рептилии. У зародышей высших позвоночных имеются четыре вида оболочек — амнион (водная), хорион (сероза), аллантоис и желточный мешок. Последний встречается у рыб, и его роль заключается в обеспечении питания зародыша. Некоторые эмбриологи не относят желточный мешок к внезародышевым оболочкам, так как в конце эмбриогенеза (по мере усвоения желтка) он втягивается в полость тела и становится частью кишечной трубки. Признаки образования амниона впервые определяются у куриного зародыша через 30 ч инкубации. Источниками развития тканей амниона являются внезародышевая эктодерма и подстилающий ее париетальный листок спланхнотома. Чтобы понять развитие внезародышевых оболочек, необходимо отметить важный морфогенетический этап эмбриогенеза — образование туловищной складки и приподнимание ранее распластанного на желтке зародыша. Таким образом зародыш несколько обособляется, но сохраняет связь с внезародышевой частью посредством узкого стебелька — пупочного шнура (канатика). Канал пупочного канатика связывает полость средней кишки зародыша с полостью желточного мешка. Все эти органы располагаются латерально от туловищной складки и относятся к внезародышевой части. Внезародышевая эктодерма и подстилающий ее париетальный листок спланхнотома формируют билатерально амниотические складки. Складки начинают обрастать зародыш с головного конца, и вскоре весь зародыш оказывается окруженным оболочкой в виде «чаши». При сближении амниотических складок справа и слева одноименные внезародышевые листки срастаются. При этом внезародышевая эктодерма оказывается обращенной к зародышу, и ее клетки дифференцируются в эпителиальную ткань с секреторной активностью. В прилежащем париетальном листке спланхнотома возникают сосуды (из спланхномезенхимы). Так создаются условия для синтеза и секреции амниотической жидкости. После замыкания амниотических складок оставшиеся части внезародышевой эктодермы и париетального листка спланхнотома формируют хорион (серозную оболочку), который обрастает зародыш. Хорион приобрел такие важные функции, как дыхательную и трофическую. В связи с содержанием материала спланхнотома в хорионе также формируются сосуды. Хорион тесно прилежит к подскорлуповой пленке на тупом конце яйца, в области воздушной камеры, из которой происходит поглощение воздуха. Как трофический аппарат хорион дает выросты, которые врастают в белковую массу. Под влиянием ферментов хориона происходит расщепление белка, и питательные вещества по сосудам хориона попадают в тело зародыша. Аллантпоис (мочевой мешок) появляется на третьи сутки инкубации в виде выроста вентральной стенки задней кишки. Он имеет колбасовидную форму и выступает из тела зародыша во внезародышевую часть яйца (экзоцелом). Стенка аллантоиса состоит из внезародышевой энтодермы, формирующей эпителиальную выстилку аллантоиса, и висцерального листка спланхнотома, мезенхима которого является источником развития сосудов аллантоиса. Постепенно аллантоис проникает в экзоцелом между хорионом и амнионом. При этом происходит срастание листков спланхнотомов аллантоиса и хориона. В связи с тем, что в составе аллантоиса рано появляются сосуды, именно они обеспечивают транспортную и дыхательную функции хориона. Однако прямая функция аллантоиса — утилизация продуктов азотистого обмена зародыша. Это происходит за счет эндоцитоза эпителиоцитами хориона экскретов зародыша и последующей секреции шлаков в полость аллантоиса. При вылуплении цыпленка тонкая ножка аллантоиса отрывается, и его дистальная часть с содержимым остается в скорлупе в виде сморщенной мембраны. Желточный мешок возникает при обрастании желтка внезародышевой энтодермой и висцеральным листком спланхнотома. Из энтодермы дифференцируется желточный эпителий, который обладает способностью расщеплять желток. Составные части последнего транспортируются по сосудистой системе, источником которой является энтомезенхима висцерального листка спланхнотома. Как отмечалось ранее, именно в стенке желточного мешка формируются первые кровеносные сосуды, здесь же локализуются стволовые клетки крови и первичные половые клетки. По мере развития зародыша объем желточного мешка уменьшается, и незадолго до вылупления он полностью втягивается в брюшную полость через пупочное отверстие и входит в состав кишки. Запаса желтка хватает на питание цыпленка в течение 4 дней после вылупления. Таким образом, в развитии внезародышевых оболочек принимают участие все три зародышевых листка, но только в той части, которая относится к внезародышевой. Маркером, определяющим границу между зародышевой и внезародышевой частями, является туловищная складка. Развитие внезародышевых оболочек у птиц происходит параллельно с развитием тела зародыша. В яйцах сумчатых желтка немного, а в ходе дробления он и вовсе отторгается, У сумчатой куницы это происходит уже в 1-м делении дробления: врезающаяся с анимального полюса борозда огибает желток сверху, так что в результате обособляются два алецитальных бластомера. У опоссума желток выталкивается из бластомер в виде отдельных гранул. Однако во всех названных случаях освободившиеся от желтка бластомеры впоследствии обрастают желтком, так что характер дробления близок к дискоидальному. Мы увидим сейчас, что то же самое можно сказать по существу и о высших млекопитающих. Развитие высших позвоночных Введение. Высшие позвоночные, к которым относятся представители класса рептилий (черепахи, ящерицы, крокодилы, змеи и др.), класс птиц и класс млекопитающих, перешли к наземному образу жизни. Развитие с метаморфозом у них заменилось прямым развитием. Естественно, что все вышеперечисленные изменения отразились на течении эмбрионального развития. К основным изменениям в ряду амфибии — рептилии — яйцекладущие млекопитающие можно отнести следующие: увеличение количества желтка в яйце (это, в свою очередь, изменило тип дробления — произошел переход от полного неравномерного дробления у амфибий к частичному дискоидальному у рептилий и птиц); изменилось строение бластулы. Развитие рептилий и птиц на суше привело к возникновению специальных приспособлений, защищающих зародыш от высыхания, механических повреждений и др. Появление толстых прочных оболочек (скорлупа, подскорлуповая, белковая) у яйца рептилий и птиц сделало невозможным наружное оплодотворение. Например, у курицы белковая оболочка и скорлупа формируются во время движения яйцеклетки по половому тракту, и оплодотворение возможно только до начала движения яйцеклетки по яйцеводу. В яйцеводе оплодотворенная яйцеклетка обволакивается секретом желез яйцевода — альбумином (это белок яйца), двумя подскорлуповыми пленками и скорлупой. Скорлупа объизвествляется, но имеет несколько тысяч пор. Сама яйцеклетка крепится к наружному слою белка вблизи тупого и острого концов с помощью двух тяжей, состоящих из уплотненного белка, — халазами (градинки). Последние удерживают яйцеклетку во взвешенном состоянии возле центра яйца. Весь путь от овуляции до откладки яйца продолжается до 26 ч. У плацентарных млекопитающих развитие зародыша стало происходить в специальном органе — матке. В связи с этим произошли изменения в строении яйцеклетки и др. Таким образом, общие признаки, характеризующие эмбриональное развитие высших позвоночных, заключаются в усложнении эмбриогенеза и появлении новых — ценогенетических признаков (преобладание внезародышевого материала над зародышевым, гетерохронность в темпах развития зародыша и внезародышевых органов). Развитие млекопитающих. Млекопитающие произошли от яйцекладущих предков — рептилий и перешли в процессе эволюции к живорождению. Эмбрион вынашивается в специальном органе — матке, где он обеспечивается питательными веществами и кислородом за счет материнского организма. Все это относится к группе плацентарных млекопитающих. Однако существуют и яйцекладущие млекопитающие и сумчатые. Они сохранили черты эмбрионального развития, свойственные птицам и рептилиям (утконос, кенгуру и др.). Развитие плацентарных млекопитающих. Органом, осуществляющим связь зародыша с материнским организмом, является плацента. Вследствие того, что в эмбриогенезе млекопитающих отпадает необходимость накопления питательного материала в яйце, возникает небольшая, диаметром 100—200 мкм вторично изолецитальная и олиголецитальная яйцеклетка. Следует отметить, что качество и количество питательного материала в яйцеклетках иное по сравнению с таковым у птиц. У птиц из содержимого яйца развиваются организм и внезародышевые органы, а содержимого яйцеклеток млекопитающих достаточно лишь для нескольких делений дробления зародыша. Оплодотворение у плацентарных млекопитающих внутреннее, и у большинства млекопитающих сперматозоиды вводятся в верхнюю треть влагалища. У некоторых, например грызунов, осеменение происходит в матке. Движению спермиев способствуют сокращения стенки матки, маточной трубы, движения ресничек эпителия маточных труб, направленный ток жидкости в маточных трубах и др. Процесс оплодотворения — достаточно сложный. Оплодотворяющая способность нативных спермиев в условиях культуры значительно ниже, чем в естественных условиях. Это связано с важной активирующей ролью на спермии органов женского полового тракта. Приобретение спермиями оплодотворяющей способности по мере их продвижения по женскому половому тракту получило название капацитации. Жизнеспособность половых клеток у большинства млекопитающих ограничена во времени. В оптимальных условиях овулировавшая половая клетка должна быть оплодотворена в течение суток. Спермии сохраняют свою оплодотворяющую способность в течение 1—2 сут, хотя подвижность их сохраняется дольше. Однако существуют примеры, когда спермии сохраняют свою оплодотворяющую способность достаточно долго, например, у летучих мышей осеменение происходит осенью, а контакт с яйцеклеткой — только весной. Слияние гамет — случайный процесс. После соприкосновения половых клеток спермий проходит через яйцевые оболочки (лучистый венец, прозрачную зону), затем плазматическую мембрану яйцеклетки, после чего становится возможным объединение генетического материала гамет. Из акросомы спермия выделяются литические ферменты, которые растворяют яйцевые оболочки. В месте контакта спермия с яйцеклеткой возникает бугорок оплодотворения. После слияния плазматических мембран спермия и яйцеклетки бугорок оплодотворения втягивается, транспортируя головку спермия в яйцо. Далее возникает оболочка оплодотворения, предотвращающая проникновение других спермиев в женскую половую клетку. Дробление зиготы полное, неравномерное, асинхронное, в результате чего вначале возникает скопление бластомеров в виде тутовой ягоды — морулы, или плотного шарика. Последующее дробление приводит к возникновению зародыша с полостью — бластоцисты. В бластоцисте выделяются две части — внутренняя клеточная масса, или эмбриобласт, и наружный слой клеток — трофобласт. Клетки трофобласта способны секретировать жидкость в полость бластоцисты и взаимодействовать со слизистой оболочкой матки, участвуя в имплантации зародыша. Следовательно, у млекопитающих в период дробления происходит раннее обособление клеток, образующих внезародышевые структуры. Это трактуется как эволюционное приобретение, связанное с внутриутробным способом развития млекопитающих. Например, у приматов в течение первых трех суток после оплодотворения зародыш продвигается по маточной трубе и к концу 4 сут имеет хорошо развитый трофобласт. Через 5 сут зародыш попадает в матку, и на 6—7-е сутки происходит его имплантация. В клетках трофобласта накапливаются протеолитические ферменты, а в клетках эпителия матки созревают рецепторы, обеспечивающие прикрепление зародыша к стенке матки. Имплантация, или внедрение, совершается в течение 2 сут и включает две фазы: прилипание (адгезия) и проникновение (инвазия). В первую фазу имплантации трофобласт дифференцируется на две части — наружную симпластическую часть, которая обеспечивает лизис цитомембран эпителиальной выстилки и подлежащих структур слизистой оболочки матки, и внутреннюю часть — клеточную (цитотрофобласт), являющуюся камбиальной частью для первой. Как только зародыш внедряется в эндометрий, трофобласт начинает выделять гонадотропный гормон, который поддерживает функционирование желтого тела в яичнике. Вокруг эмбриобласта рано образуется внезародышевая мезенхима, подстилающая трофобласт изнутри. Обе структуры ответственны за развитие хориона — ворсинчатой оболочки, которая с участком слизистой оболочки матки формирует плаценту (детское место). Различают несколько видов плацент. Эпитпелиохориальная (полуплацента) — хорион зародыша прилегает к эпителию слизистой оболочки матки, а ворсинки хориона проникают в углубления слизистой оболочки матки, не нарушая ее целостности. Питательные вещества и кислород из сосудов стенки матки поступают в кровеносные сосуды ворсинок хориона и далее — в зародыш. Данный вид плаценты встречается у свиньи, лошади, китообразных. Десмохориальный вид плаценты встречается у жвачных животных. В этом случае ворсинки хориона разрушают эпителий матки и внедряются в соединительнотканную строму слизистой оболочки. В эндотелиохориальной плаценте формируется еще более тесная связь сосудов ворсинок хориона с тканями матки — ворсинки хориона достигают стенки маточных сосудов вплоть до эндотелия (этот тип плаценты характерен для хищных животных). Гемохориалъная плацента — при ее формировании ворсинки хориона разрушают эндотелий сосудов и омываются кровью материнского организма (приматы, человек). Таким образом, в развитии млекопитающих наблюдается важное эволюционное изменение характера развития зародыша и его временных органов. Вследствие асинхронного дробления бластомеров очень рано и быстро формируется трофобласт — первый внезародышевый орган, обеспечивающий прикрепление зародыша к стенке матки и его питание. Обособление внезародышевой мезенхимы обеспечивает раннее развитие кровеносной системы и элементов крови. Только после имплантации зародыша становятся возможными развитие других внезародышевых органов и переход к гаструляции, гисто- и органогенезу. Развитие желточного мешка, хотя и происходит, функцию питания зародыша он не выполняет. В стенке желточного мешка формируются первичные половые клетки, стволовые клетки крови, а также сосуды. При этом амнион, аллантоис и желточный мешок развиваются из клеток эмбриобласта. Амнион появляется до возникновения первичной полоски. У разных млекопитающих механизм образования амниона разный — от видимого сходства с развитием амниона у птиц до возникновения амниотического пузырька в составе эмбриобласта (или сочетания двух поименованных способов). С возникновением амниотического пузырька оставшийся материал эмбриобласта расщепляется способом деламинации на верхнюю (эпибласт) и нижнюю (гипобласт) клеточные массы. Эпибласт при этом оказывается дном амниотического пузырька. Клетки гипобласта далее выстилают первичный желточный мешок, который затем разделяется перетяжкой на две части — кишечную трубку и собственно желточный мешок. Короткий вырост хвостовой части кишки представляет собой аллантоис. Последний, как и желточный мешок, не выполняет функции мочевого мешка, однако сосуды аллантоиса имеют большое значение в жизнеобеспечении зародыша. При соприкосновении стенки аллантоиса с внутренней поверхностью хориона улучшается кровоснабжение и питание зародыша. После выделения внезародышевых органов, которые могут обеспечить продолжение развития зародыша, заметно активизируются пролиферативные и миграционные процессы в эпибласте. Они практически аналогичные тем, которые происходят в эпибласте куриного зародыша. Формируются первичная полоска и гензеновский узелок, клетки которых закономерно мигрируют в пространство между эпи- и гипобластом. Таким образом формируется многослойный зародыш, в составе которого далее выделяются эктодерма с нервной пластинкой, хордомезодерма и энтодерма (в головную часть последней встраивается материал прехордальной пластинки). В последующем происходят сегментация и нейруляция. которые, как было показано на примере развития низших позвоночных, происходят под индуцирующим влиянием хорды. Зародыш связывается с плацентой посредством пупочного канатика. Плацента — губчатое образование, состоящее из кровеносных сосудов зародыша, окруженных стромой и клетками хориона (цито- и симпластотрофобласта). В состав плаценты также входит так называемая материнская часть, состоящая из видоизмененной стенки матки с ее тканевыми и органными элементами. В стенке матки зародыш находится интрамурально и заключен в амнион. Продолжительность эмбриогенеза у различных млекопитающих колеблется в широких пределах — от 21 сут. (у крыс), 4 нед. (у кролика), 7—8 мес. (у человекообразных обезьян) до 18—22 мес. (у слонов). Рождение зародыша происходит в результате ритмических сокращений стенки матки, разрыва амниона и выхода зародыша во внешнюю среду (при этом пуповина перегрызается самкой). В последующем рождается и плацента (в связи с чем ее именуют последом).
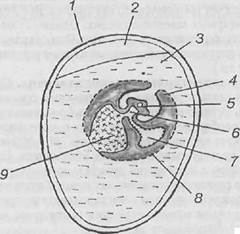
Рис. Поперечные разрезы зародышей ланцетника (по: Конклинуи Гатчеку, из Кнорре, 1967): А - карта презумптивных зачатков яйцеклетки ланцетника; Б - поперечный срез бластулы ланцетника; В, Г - поздняя гаструла (продольный и поперечный разрезы); Д - превращение нервной пластинки, погрузившейся под кожную эктодерму, в нервный желобок и начало обособления мезодермальных карманов от первичной кишки; Е - обособление замкнутых целомических мешков, замыкание энтодермы в кишечную трубку и формирование хордального тяжа. Б; 1 - крыша бластулы, 2 - дно бластулы, 3 - краевая зона бластулы; В: 4 - дорсальная губа бластопора, 5 - мелкоклеточный материал вентрального серпа,
Рис. Схема развития внезародышевых органов у птиц (Корниг, Заварзин, из Кнорре, 1967): 7 - скорлупа, 2 - воздушная камера, 3 - белочная оболочка (белок), 4 - амниотическая складка, 5 -зародыш, 6- кишечная трубка, 7 - аллантоис, 8- формирование серозной оболочки (хориона), 9 - обрастание желтка и образование желточного мешка
Рис. Схема последовательных стадий образования внезародышевых органов у млекопитающих (Заварзин, из Кнорре, 1967): 7 - зародыш, 2 - туловищная складка, 3 - амниотические складки, 4 - амнион, 5 - желточный мешок, 6 - аллантоис, У – хорион
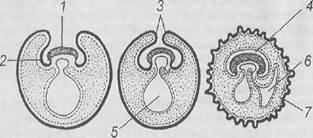
Рис. Схема дробления и образования бластоцисты млекопитающих (Дюваль, Ван Бенеден, Заварзин, из Кнорре, 1967): А - полное асинхронное дробление зиготы, стадии 2 и 4 бластомеров, Б – обрастание светлыми клетками темных (морула), В - образование бластоцисты, формирование полости, обособление темных клеток внутренней клеточной массы от светлых - наружной клеточной массы - трофобласта; 1 - внутренняя клеточная масса, 2 – трофобласт
Рис. Схема голобластического чередующегося дробления (а) и первое деление дробления (б), стадия двухклеточного зародыша (Гертиги Рокк, из Гистология, 1972): а: 1 - меридиальное; 2 - широтное деление бластомера; б: 1 - прозрачная зона; 2 - бластомеры, 3 - ядра бластомеров
|
||||
|
|
Последнее изменение этой страницы: 2016-06-22; просмотров: 812; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 52.15.170.196 (0.018 с.) |