Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Лекция №6. Мезодерма и ее производные.Содержание книги
Поиск на нашем сайте
Мезодерма, с которой связано развитие всех органов, состоит из парных частей: дорсальная, осевая разделяется на сомиты; латеральная, несегментированная - боковая пластинка или спланхнотом (греч.: splanchnon - внутренности, tomos - отрезок). Принято различать два принципиально отличных типа закладки мезодермы. Первый — телобластический — встречается в наиболее чистом виде у спирально дробящихся форм. Мы уже упоминали о бластомерах 2d и 4d, получивших в ходе делений дробления всю полярную плазму. Две крупные клетки потомства бластомера 4d, симметрично расположенные в полости бластоцеля в области губ бластопора, дают начало всей так называемой целомической мезодерме личинки. Эти бластомеры называются мезобластами, или мезотелобластами. Более мелкие мезодермальные клетки отпочковываются от этих крупных бластомеров путем серии последовательных делений. В результате возникает пара так называемых мезодермальных полосок. Позже они подразделяются на парные отдельности — сомиты, внутри которых путем расхождения клеток образуются участки вторичной полости тела, или целома. Способ образования полостей путем расхождения клеток называется шизоцельным, или кавитационным. Таким образом, при телобластическом способе закладки целомическая мезодерма образуется из двух бластомеров со строго определенной генеалогией. Мезодерма при этом никак не связана с энтодермой, образующейся из других бластомеров. Закладка мезодермы из отдельных предназначенных к тому бластомеров имеет место также у большинства круглых червей, некоторых ракообразных и в ряде других групп первичноротых животных. В разных систематических группах генеалогия порождающих мезодерму бластомеров весьма различна. Принципиально другой — энтероцельный — способ закладки мезодермы наблюдается в наиболее ясной форме у иглокожих, ланцетника, кишечнодышащих, плеченогих. Здесь материал будущей мезодермы вворачивается вместе с энтодермой в составе единого гастрального впячивания, и в процессе инвагинации граница между обеими закладками, как правило, неразличима. Только прослеживая судьбу закладок в ретроспективном порядке, т. е. идя от поздних стадий развития назад, к ранним, можно выяснить, какую часть гастрального впячивания выстилает материал будущей мезодермы. Такое впячивание, включающее в себя материал и энтодермы и мезодермы (а у хордовых еще и хорды), называется первичным кишечником, или архентероном. Соответственно гастроцель в этих случаях называется полостью первичной кишки, или полостью архентерона. Мезодерма выделяется из архентерона путем выпячивания его стенок и отшнуровки возникших выпячиваний, реже — путем деламинации стенок архентерона или иммиграции клеток из них. После отделения мезодермы в составе стенки архентерона остается уже чисто энтодермальный материал, и архентерон превращается в полость вторичной (дефинитивной) кишки. Как и полость сомитов первичноротых, полость отшнуровавшихся мезодермальных пузырьков (часть бывшей полости архентерона) называется целомом. Дальнейшую дифференцировку мезодермы мы рассмотрим ниже. Уже говорилось, что телобластический и энтероцельный способы в чистом виде встречаются у сравнительно немногих форм. Но эти формы относятся к двум разным ветвям животного мира — к первично- и вторичноротым животным. Как известно из курса зоологии, первичноротыми называют животных, у которых отверстие бластопора непосредственно превращается в ротовое отверстие, а вторичноротыми — у которых ротовое отверстие закладывается вторично, на стороне тела, противоположном бластопору (бластопор же нередко превращается в анальное отверстие). «Сердцевину» ствола первичноротых образует группа Spiralia с телобластической закладкой мезодермы. Огромный тип членистоногих, у которых телобластичность почти утрачена, естественно выводится из типичных Spiralia — кольчатых червей. С другой стороны, у основания ствола вторичноротых следует поставить иглокожих с ярко выраженной энтероцельностью. Хордовые, у большинства которых энтероцсльность затушевана, несомненно, относятся к тому же стволу. Развитие производных мезодермы Осевая мезодерма, У всех позвоночных имеются осевая и боковая мезодермы, причем осевая мезодерма подразделяется на сомиты (метамеризуется). Способ закладки и дифференцировки сомитов в разных классах хордовых неодинаков. У ланцетника сомиты формируются в виде энтероцельных выпячиваний архентерона и с самого начала содержат участок целомической полости. У большинства позвоночных сомиты сначала закладываются в виде сплошных скоплений мезодермальных клеток и лишь позже в них возникают полости путем расхождения этих клеток. В ходе дальнейшего развития сомита из его клеток образуются три основные закладки. Наружная, обращенная к эктодерме стенка сомита формирует кожный листок, или дерматом. Из его клеток впоследствии возникает соединительная часть кожи, представленная преимущественно фибробластами. Внутренняя часть сомита, примыкающая к хорде (низшие позвоночные) или к хорде и нервной трубке (высшие позвоночные), образует склеротом — зачаток осевого скелета, вскоре распадающийся на отдельные клетки. Часть сомита, расположенная между дерматомом и склеротомом,— миотом — зачаток всей поперечно-полосатой мускулатуры. В разных классах позвоночных соотношение и темпы развития этих частей сомита неодинаковы. У низших позвоночных основная часть сомитов, как правило, представляет собой миотомы. У высших позвоночных сомиты вначале подразделяются на дерматом и массу склеротомпых клеток, а миотом (точнее, скопление эмбриональных мышечных клеток—миобластов) появляется позже на внутренней поверхности дерматома. Некоторые авторы полагают, что миобласты возникают путем размножения клеток дерматома, другие — что они перемещаются на его поверхность из более внутренних областей, отделяясь от массы склеротомальных клеток. Вначале осевая мезодерма метамеризуется не только в туловищной, но и в головной части тела зародыша, Однако во взрослом состоянии лишь у ланцетника в области головы сохраняется метамерная структура. У других позвоночных головные сомиты распадаются вскоре после своего возникновения. Основная часть их клеток образует парные хрящевые закладки задней части черепа — парахордалии. Таким образом, эта клеточная масса по своим потенциям соответствует склеротомам. Передние концы парахордалий, как и передний конец хорды, находятся на уровне вентральной мозговой складки. Спереди от нее возникают еще две парные Г-образные хрящевые закладки черепа — трабекулы. Их задняя часть строится из мезенхимы прехордальной пластинки, а передняя — из клеток нервного гребня (как и висцеральный скелет). Туловищные сомиты всех позвоночных в конце концов также распадаются, но намеченная ими метамерия тела у взрослых животных сохраняется. Во-первых, это связано с тем, что сомиты определяют расположение спинномозговых (спинальных) нервных ганглиев, во-вторых, выходящие из спинальных ганглиев нервные окончания прорастают всегда через передние, задние половины сомитов, даже если сомит повернуть на 180° относительно оси тела зародыша (в последнем случае нервные окончания будут прорастать по-прежнему через те части сомитов, которые исходно были направлены вперед, а теперь — назад). В-третьих, метамеризация закрепляется в расположении тел позвонков: каждый позвонок возникает из передней части более заднего сомита и задней части более переднего сомита. Развитие органов выделения. У анамний последовательно развиваются два сменяющих друг друга органа выделения: головная почка, или предпочка (пронефрос), и туловищная, или первичная, почка (мезонефрос). У взрослых анамний функционирует обычно мезонсфрос, хотя у личинок и даже у взрослых круглоротых и некоторых костистых рыб пронефрос также участвует в функции выделения. У амниот вслед за пронефросом и мезонефросом развиваются расположенная каудальнее тазовая почка — метанефрос, которая и функционирует (хотя у сумчатых млекопитающих до достижения половой зрелости действует мезонефрос). Все три типа почек образуются из мезодермы, находящейся в области ножек сомитов. Пронефрос развивается из ножек немногих передних сомитов, мезонефрос — из ножек почти всех туловищных сомитов, а метанефрос—из расположенного каудальнее скопления нефрогенной мезенхимы. Наиболее четко метамеризация выражена в развитии пронефроса. Стенки его канальцев образуются непосредственно из стенок сомитных ножек. Поэтому канальцы пронефроса открываются своими внутренними концами в полость целома. Эти концы имеют вид воронок, покрытых ресничками,— нефростомов. Противоположные концы канальцев загибаются назад и сливаются друг с другом в парные продольные тяжи, из которых развиваются вольфовы каналы, или первичные мочеточники. Вольфовы каналы продолжают расти назад, индуцируя образование мезонефрических канальцев в более задних сегментах тела. Канальцы мезонефроса также возникают из мезодермы сомитных ножек, но у большинства позвоночных ко времени образования мезонефрических канальцев мезодерма ножек отшнуровывается от сомитов и преобразуется в мезенхимную ткань. Из этой ткани и формируются метамерные мезонефрические канальцы, у которых впоследствии появляются многочисленные изгибы и ответвления. У зародышей анамний внутренние концы мезонефрических канальцев вторично соединяются с целомом посредством ресничных воронок, а у высших позвоночных канальцы слепо заканчиваются в мезенхиме. Наружные концы; канальцев открываются в первичный мочеточник, который индуцирует само образование канальцев. У высших позвоночных от мезонефроса остаются лишь небольшие придатки половых желез — эпоофорон у самок и эпидимис у самцов. Функционирующей почкой у высших позвоночных, как уже говорилось, является метанефрос (тазовая почка). В строении метанефроса не остается уже никаких следов метамерии, и он не связан с целомом ни на одной стадии развития. Тем не менее нефрогенная мезенхима, из которой метанефрос построен, произошла из того же источника, что и материал про- и мезонефроса,— из ножек сомитов. Важную роль в развитии тазовой почки играет первичный мочеточник. От него к скоплению нефрогенной мезенхимы растет отросток с расширенным концом. Сам отросток превращается во вторичный мочеточник, а его расширенный конец — в почечную лоханку. На поверхности лоханки также образуются выпячивания, из которых развиваются верхние отделы выводных путей почки. Позже они открываются в мочевые канальцы, которые образуются уже из нефрогенной мезенхимы, но под индукционным воздействием вторичного мочеточника. Мочевые канальцы тесно соприкасаются с клубочками кровеносных капилляров, образуя вместе с ними мальпигиевы тельца — органы фильтрации высших позвоночных. Половые железы и половые протоки. Стенки половых желез позвоночных развиваются из висцерального листка боковой пластинки на уровне ножек сомитов. Эти утолщения получили не совсем удачное название герминативного эпителия. Недостаток этого укоренившегося термина состоит в том, что он как бы подразумевает происхождение половых клеток из этого эпителия. На самом деле половые клетки возникают из первичных гоноцитов и лишь позже заселяют половые железы. Герминативный эпителий — это соматическая ткань, образующая стенку половой железы. Сама железа па ранних стадиях своего развития представляет собой складку, вдающуюся в полость тела,— так называемую половую складку. Эта складка постепенно заполняется окружающей мезенхимой, за счет которой развивается внутренняя (мозговая) часть железы. До определенной стадии развития железа имеет одинаковое для обоих полов строение. Затем под влиянием проникших в нее первичных половых клеток, а также в зависимости от гормонального баланса организма железа дифференцируется либо в семенник, либо в яичник. Для яичника характерно преимущественное развитие корковой части (из которой впоследствии образуется фоликулярный эпителий, окружающий ооциты), для семенника — мозгового слоя. Неодинаково в зародышах разного пола идет и развитие выводных протоков половых желез. У самцов семенные канальцы, где происходит сперматогенез, соединяются с вольфовыми каналами, которые принимают на себя функции семяпроводов. У амниот вынос семени — единственная функция вольфовых каналов, так как связанный с тазовой почкой вторичный мочеточник развивается из специального выроста вольфова канала. У анамий, где функционирующей почкой является мезонефрос, вольфовы каналы объединяют функции мочеточника и семяпровода. В эмбриогенезе позвоночных появляется еще одна пара каналов, идущих параллельно вольфовым,— мюллеровы каналы. У самцов они позже дегенерируют, а у самок сохраняются и превращаются в яйцеводы. У многих анамний по крайней мере верхние отделы мюллеровых каналов развиваются за счет клеток резорбирующего пронефроса. Поэтому эти каналы открываются в полость тела (целом) одним из нефростомов пронефроса, превратившегося в воронку яйцевода (это хороший пример субституции — замещения функций, что, по мнению ряда авторов, является одним из главных путей эволюции органов). При овуляции яйцо выходит сначала в полость тела и уже затем захватывается воронкой яйцевода (мюллерова канала). Производные боковой пластинки. Расположенная вентральнее ножек сомитов боковая пластинка очень рано разделяется на париетальный и висцеральный листки. Между ними находится вторичная полость тела (целом), и оба листка образуют ее выстилку. Соответственно своему положению париетальный листок формирует внешнюю выстилку целома, а висцеральный — внутреннюю. Оба листка соединяются друг с другом по средней линии тела посредством спинной и брюшной брыжеек. Рассмотрим сначала развитие производных висцерального листка. К ним относят сердце, кровеносные сосуды и клетки крови. Кроме того, по данным П. Ньюкупа, у хвостатых амфибии из висцерального листка образуются первичные половые клетки (гоноциты). Все эти закладки для полноценного развития нуждаются в контакте с энтодермой. Так, презумптивная кроветворная мезодерма не формирует кровяных островков в отсутствие контакта с энтодермой, но наличие таких контактов ускоряет этот процесс. Развитие сердца. У птиц парный зачаток сердца возникает в середине вторых суток инкубации в виде двух симметрично расположенных утолщений висцерального листка мезодермы, который тесно связан с энтодермой. Левый и правый зачатки соединяются лишь после сворачивания энтобласта в трубку головной кишки, причем вентральнее последней. Из объединившихся трубок висцеральной мезодермы возникает мышечная стенка сердца — миокард. Внутренняя оболочка сердца— эндокард — также получается в результате слияния двух трубчатых зачатков, образованных мигрировавшими по энтобласту и миокарду мезенхимными клетками. Единая сердечная трубка переходит в широкие желточные вены, несущие кровь от внезародышевой системы кровообращения со стенки желточного мешка. Сердечная трубка лежит в широкой перикардиальной полости, являющейся частью целома. Точно так же, как у цыпленка, развивается сердце у всех других амниот. В отношении цитодифференцировки сердечная мышца отличается от скелетной тем, что здесь не сливаются миобласты и не образуются мышечные волокна. На протяжении всего гистогенеза эта ткань сохраняет клеточное строение. Кровеносные сосуды позвоночных развиваются, по-видимому, исключительно из мезенхимы. Они закладываются в виде не связанных друг с другом кровяных островков — клеточных скоплений, внутри которых позже образуются просветы. Затем отдельные трубочки сливаются в рыхлую сеть. Эти стадии развития особенно хорошо видны на краю бластодиска зародышей птиц, но по существу не отличаются и у других позвоночных. Наружные клетки островков (ангиобласт) уплощаются и вступают в контакт друг с другом, образуя эндотелиальную стенку сосуда, а внутренние клетки (гемобласт) превращаются в клетки крови. Первые крупные сосуды зародыша — парные желточные вены, впадающие в трубчатый зачаток сердца сзади и несущие (у амниот) к сердцу кровь от внезародышевых частей, а также выходящий из переднего конца зачатка сердца ствол аорты, разделяющийся на два артериальных ствола. Расположение возникающих в дальнейшем кровеносных сосудов в основном определяется окружающими их морфологическими структурами. Так, в головной области зародышей всех позвоночных вначале образуются 6 парных дуг аорты — по числу жаберных дуг. У высших позвоночных большинство этих сосудов впоследствии дегенерирует. Вообще в начале развития возникает избыточное количество мелких сосудов, часть которых в дальнейшем запустевает или превращается в капилляры. Лишь те сосуды, направление которых соответствует анатомическим особенностям тела взрослого животного и через которые проходит достаточно мощный кровяной поток, превращаются в развитые кровеносные стволы. Именно изучая развитие кровеносной системы, В. Ру впервые пришел к принципу эквифинальности: один и тот же план строения кровеносной системы взрослого организма создается разными путями из беспорядочных, вариабельных «узоров» эмбриональных кровеносных сосудов. Развитие парных конечностей. Парные конечности позвоночных развиваются из мезенхимных клеток, выселившихся из париетального листка мезодермы и покровной эктодермы. У зародышей амфибий ранние зачатки конечностей имеют вид обособленных бугорков У зародышей амниот вначале формируются длинные складки, растянутые в передне-заднем направлении (вольфовы гребни), которые позже рассасываются в своей средней части; из их передних и задних концов развиваются соответственно передние и задние конечности. На самых ранних стадиях роста конечностей их эктодермальный эпителий пассивно растягивается размножающейся мезенхимой; вскоре и эктодерма начинает активно участвовать в росте конечности. У амниот эктодерма верхушки конечности утолщается, образуя так называемый апикальный гребешок. По мере роста конечности меняется ее форма: апикальная часть расширяется и уплощается, зачаток конечности скручивается вокруг своей длинной оси. На апикальной поверхности появляются зачатки пальцев. У амниот обособление пальцев связано с гибелью клеток в промежутках между их зачатками. Одновременно с внешней дифференцировкой конечности формируется ее внутренний скелет путем образования хрящей из сгущений мезенхимных клеток. Первым выделяется зачаток проксимального хряща — стилоподия, из которого в передней конечности разовьется плечевая кость, а в задней — бедренная. Затем образуются хрящи следующей в дистальном направлении части — зигоподня (локтевой и лучевой хрящи в передней конечности, большой и малый берцовые — в задней) и, наконец, аутоподия (хрящи кисти или стопы и фаланг пальцев). Хрящи плечевого и тазового поясов формируются позже стилоподиев, но раньше аутоподиев. В конечность прорастают кровеносные сосуды и миобласты из сомитов. При дифференцировке парных конечностей происходят интенсивные эпителиально-мезодермальные взаимодействия. На начальных стадиях развития конечности, по-видимому, основным является воздействие мезодермы на эктодермальный эпителий. Под влиянием мезодермы эпителий утолщается и начинает активно расти. В дальнейшем нормальную дифференцировку дистальных отделов конечности (образование пальцев) определяют обратные влияния, исходящие от утолщенного эпителия верхушки почки конечности (уже упоминавшегося ранее апикального гребешка), на мезодерму конечности. При удалении апикального гребешка фаланги не дифференцируются, а при его пересадке на презумптивную мезодерму проксимальной части конечности (из которой в норме должны были бы дифференцироваться бедренный или плечевой отделы) из нее развиваются дистальные части конечности — плюсна (или кисть) и фаланги. Интересно, что проксимальная мезодерма конечности не пассивно «прочитывает» сигналы, исходящие из гребешка, а как бы интерпретирует их «по-своему»: если мезодерму проксимальной части задней конечности (ноги) зародыша курицы пересадить под гребешок передней конечности (крыла), то она образует дистальную часть, но не крыла, а задней конечности. Значит, в «интерпретации» индукционного воздействия определенную роль сыграла природа самого реагирующего материала, взятого от задней конечности. Другая морфогенетически активная зона зачатка конечности — небольшая область на ее заднем крае, около основания. Если эту так называемую «зону поляризующей активности» пересадить на передний край конечности, то произойдет ее зеркальное удвоение: спереди появится второй задний кран с соответствующими пальцами. Если же эту зону удалить — конечность станет симметричной, задне-передние различия в ее структуре исчезнут. Обнаружено, что зона поляризующей активности характеризуется повышенной концентрацией морфогенетически активного вещества — ретиновой кислоты (производное витамина А).
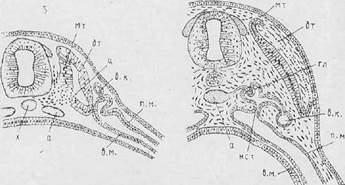
Рис. Четыре последовательные стадии (А—Г) — развития производных мезодермы (по А. А. Заварзину): а — аорта; вк— вольфов канал; вм — висцеральный листок мезодермы; гл —гломус; дт — дерматом; мт — миотом; нет—нефростом; нт — нефротом; пм—париетальный листок мезодермы; X — хорда; ц — целом
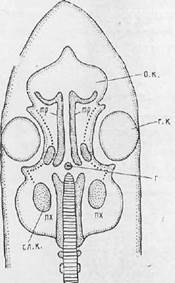
Рис. Схема расположения закладок черепа у зародыша позвоночного: г— гипофиз; гк — глазная капсула; ок —обонятельная капсула; пх —парахордалин; сл.к. — слуховая капсула; тр — трабекула
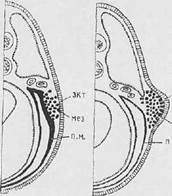
Рис. Последовательные стадии (А, Б) развития почки конечности земноводных: мез — мезенхима почки конечности; пм — париетальный листок мезодермы; экт – эктодерма.
Рас. Зародыши амфибий на стадии раннем хвостовой почки в сагиттальной проекции: А — вид с удаленной покровной эктодермой; Б—вид разреза в сагиттальной плоскости; ан — анальное отверстие; сч.п. — слуховой пузырек; гл.п, — глазной пузырь; жаб.щ. — жаберные щели; ж.энт. — желточная энтодерма; зач.пш. — зачаток гипофиза; з.к. — задняя кишка; м — мозг; мезод —мезодерма; Н-тр. — нервная трубка; п.м. — передний мозг; п.к. — передняя кишка; печ.в.— печеночный вырост; р — место будущего рта; пр. — пронефрос; с — сердце; сом— сомит; ср.к.— средняя кишка; х — хорда; жаб.д. — жаберные дуги; ср.м.—средним мозг
Рис.. Развитие органов пищеварения и их производных у амфибий; А— фронтальный; Б — сагиттальный разрез зародыша на стадии ранней хвостовой почки; В—схема строения передней части кишечника на более поздней стадии развития (сагиттальная проекция); гл — глотка; гл.к. — глоточные карманы; дв.к. —- двенадцатиперстная кишка; ж — желудок; желчн.п. — желчный пузырь; л — зачаток легкого; п — зачаток печени; печ.в. — печеночный вырост; п.пк, —полость передней кишки; ср.к. — средняя кишка; х —хорда
|
|||||
|
|
Последнее изменение этой страницы: 2016-06-22; просмотров: 2082; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.224.30 (0.011 с.) |