
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Проецирование световых лучей на сетчатку глазаСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте Прежде чем попасть на сетчатку, световые лучи последовательно проходят через роговицу, жидкость передней камеры глаза, хрусталик и стекловидное тело, вместе образующие оптическую систему глаза (рис. 17.5). На каждом из этапов этого пути свет преломляется и в результате на сетчатке возникает уменьшенное и перевернутое изображение наблюдаемого предмета, этот процесс называется рефракцией. Преломляющая сила оптической системы глаза составляет около 58,6 диоптрий при рассматривании удаленных предметов и возрастает до приблизительно 70,5 диоптрий при фокусировании на сетчатку световых лучей, отраженных от близко расположенных предметов (1 диоптрия соответствует преломляющей силе линзы с фокусным расстоянием 1 м). 17.3.1.1. Аккомодация Чтобы фокусировать на сетчатке световые лучи, отраженные от близко расположенных предметов, оптическая система глаза должна преломлять их тем сильней, чем ближе расположен наблюдаемый объект. Механизм, с помощью которого глаз настраивается на рассмотрение удаленных или
Склера Сосудистая \ оболочка
эпителий Рис. 17.5. Оптическая система глаза и проекция световых лучей на сетчатку. Световые лучи, отраженные от рассматриваемой части наблюдаемого объекта (точка фиксации), преломляются оптическими средами глаза (роговица, передняя камера, хрусталик, стекловидное тело) и фокусируются в центральной ямке сетчатки. Проекция световых лучей на поверхность центральной ямки обеспечивает максимальную остроту зрения благодаря малым размерам рецептивных полей и отсутствию ганглиозных и биполярных клеток на пути прохождения световых лучей к фоторецепторам. 77. Сенсорные системы • 719
При максимальном сокращении ресничных мышц кривизна хрусталика становится максимальной, что позволяет фокусировать на сетчатке световые лучи, отраженные от ближайшей точки ясного видения. У детей эта точка расположена примерно в семи сантиметрах от глаза, но с возрастом эластичность хрусталика уменьшается, что ограничивает возможность увеличения его кривизны и соответственно преломляющей силы. В связи с возрастным уменьшением диапазона аккомодации (интервала изменения преломляющей силы хрусталика) ближайшая точка ясного видения постепенно удаляется: в 20 лет она располагается на расстоянии около 10 см от глаза, в 30 лет — 14 см, в 40 лет — 22 см, в 50 лет — 40 см и т. д. Возрастное уменьшение аккомодационной способности (пресбиопия, или возрастная дальнозоркость) принуждает человека использовать при чтении очки с двояковыпуклыми линзами. 17.3.1.2. Аномалии рефракции У большинства людей глаз имеет шарообразную форму, максимально приспособленную для того, чтобы световые лучи фокусировались на сетчатке при любой преломляющей силе хрусталика, изменяющейся в процессе аккомодации. При нормальной рефракции на сетчатке одинаково четко проецируются изображения далеких и близких предметов, а проявление такой универсальной рефракционной способности определяют термином эмме-тропия. Аномалии рефракции возникают при изменениях формы глаза, проявляющихся в удлинении его оптической оси (близорукость, или миопия) или в ее укорочении (дальнозоркость, или гиперметропия). При удлиненной форме глаза, характерной для миопии, обычная преломляющая способность его оптической системы оказывается избыточной, в связи с чем изображение фокусируется перед сетчаткой и субъективно воспринимается нечетким. Дефект зрительного восприятия ощущается только при рассматривании удаленных предметов, тогда как световые лучи, отраженные от близко находящихся предметов, фокусируются точно на сетчатке благодаря ограничению механизма аккомодации. Например, при чтении близоруким людям нужна меньшая, чем людям с нормальной рефракцией, преломляющая сила хрусталика, поэтому нагрузка на ресничные мышцы у них снижается. Но для ясного видения далеких предметов близорукому человеку приходится носить очки с двояковогнутыми рассеивающими линзами, уменьшающими преломление световых лучей. При характерной для гиперметропии укороченной оптической оси глаза световые лучи фокусируются за сетчаткой, что принуждает ресничные мышцы к сокращению даже при рассмотрении далеких предметов. Для компенсации 720 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Наибольшей преломляющей способностью в оптической системе глаза обладает роговица (около 48,8 диоптрий), которая не участвует в аккомодации. Поверхность роговицы не идеально сферична почти у всех людей, ее кривизна по вертикали выражена больше, чем по горизонтали. При значительном отклонении формы роговицы от идеальной пропорции возникает аномалия рефракции, которая определяется как астигматизм. В результате такой аномалии световые лучи, отраженные от наблюдаемых объектов, преломляются не одинаково, поэтому часть лучей фокусируется на сетчатке, а остальные — за ее пределами, что ведет к искажению зрительного восприятия. Для исправления этого дефекта применяются индивидуально подобранные линзы очков с различной кривизной по вертикальному и горизонтальному меридианам. 17.3.1.3. Регуляция интенсивности светового потока Приспособление к изменяющимся условиям освещения происходит с помощью зрачкового рефлекса. Ширина зрачка уменьшается при ярком свете благодаря сокращению кольцевых мышц, управляемых парасимпатическими волокнами глазодвигательного нерва, а при слабом освещении зрачок расширяется с помощью радиальных мышц, получающих симпатическую иннервацию. Уменьшая просвет зрачка, глаз защищается от избытка света, а увеличивая ширину зрачка, он повышает чувствительность зрительной системы к воспринимаемым стимулам. Сужение зрачков повышает глубину резкости, что позволяет лучше видеть удаленные предметы. При расширении зрачков глубина резкости снижается, а вместе с ней снижается острота зрения, которая характеризуется максимальной способностью глаза различать две соседние точки зрительного пространства как отдельные. В норме глаз различает две точки, видимые под углом в одну минуту при достаточно ярком освещении. 17.3.1.4. Проекция зрительного поля на сетчатку Благодаря механизму аккомодации глаз представляет собой самофокусирующуюся оптическую систему, позволяющую получать на сетчатке четкое изображение приближенных и удаленных предметов. Кроме того, глаз подвижен, что позволяет ему следить за перемещающимися объектами. Движения глаза обеспечиваются сокращениями глазодвигательных мышц (двух косых и четырех прямых), которые позволяют повернуть глаз внутрь или наружу, вверх или вниз. Оба глаза поворачиваются бинокулярно, т. е. содружественно, и при рассмотрении близких предметов глазные оси сходятся {конвергенция глаз), а при рассмотрении далеких предметов — расходятся {дивергенция). Благодаря бинокулярному зрению каждая точка зрительного поля проецируется на соответствующие друг другу участки сетчатки обоих глаз. Изображение одного и того же предмета на сетчатках левого и правого глаза немного различается, поскольку глаза занимают разную позицию по отношению к зрительному полю. Разница изображений наблюдаемого предмета на сетчатке левого и правого глаза обозначается термином поперечная диспарация, она способствует последующему преобразованию двухмерного изображения на сетчатке в трехмерное изображение, создаваемое в процессе переработки зрительной информации в коре. 17. Сенсорные системы • 721
Рис. 17.6. Ретинотопическая проекция сетчатки в первичную зрительную кору. Пространственное соотношение между объектами зрительного поля соблюдается в его проекции на соответствующие друг другу (корреспондирующие) области сетчатки каждого глаза. Афферентные сигналы от носовых половин сетчатки благодаря частичному перекресту волокон зрительных нервов поступают в противоположное полушарие. Афферентные сигналы от височных половин сетчаток попадают с помощью неперекрещивающихся волокон зрительных нервов в ипсилатеральное полушарие. В связи с этим в каждое полушарие поступает зрительная информация о противоположной ему половине зрительного поля и с сохранением его пространственных координат (принцип ретинотопии). При направленном на определенные объекты внимании световые лучи, отраженные от них, всегда фокусируются в центральной ямке. Поэтому центральная ямка и прилегающие к ней области сетчатки имеют наибольшую площадь представительства в первичной зрительной коре. Зрительное поле представляет собой область, видимую обоими глазами в тот момент, когда они и голова неподвижны. При таком условии левая половина зрительного поля проецируется на носовую половину сетчатки левого глаза и височную половину сетчатки правого глаза. Соответственно, правая половина зрительного поля проецируется на носовую половину сетчатки правого глаза и височную половину сетчатки левого глаза. Поскольку оптическая система глаза перевертывает изображение, верхняя половина зрительного поля проецируется на нижнюю половину сетчатки, а нижняя половина зрительного поля — на верхнюю половину сетчатки. При дальнейшей переработке информации в наружном коленчатом теле и первичной зрительной коре сохраняются все пространственные координаты зрительного поля: каждая область сетчатки связана с соответствующим представительством в коре, что называется ретинотопией (рис. 17.6). 722 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
При наблюдении за движущимися в зрительном поле объектами, а также при движении человека относительно окружающего мира происходят следящие движения глаз, благодаря которым изображение в одной и той же области сетчатки сохраняется неизменным. При зрительном восприятии неподвижных объектов, имеющих многочисленные детали формы, а также во время чтения происходят быстрые движения глаз, предназначенные для фиксации наиболее информативных деталей объекта. Например, при восприятии человеческого лица фиксируются его контуры, но с наибольшим вниманием рассматриваются глаза и рот, изображения которых глаз проецирует не вообще на сетчатку, а на ту ее область, которая обладает максимальной разрешающей способностью. Это центральная ямка, или /оуео, представляющая собой небольшое углубление диаметром около 3 мм в центре сетчатки. При рассматривании любых объектов глаза ежесекундно совершают около трех очень быстрых непроизвольных и субъективно не ощущаемых движений, которые называются саккадами. Благодаря таким движениям изображение на сетчатке регулярно смещается, вызывая раздражение разных фоторецепторов. Необходимость саккад объясняется свойством зрительной системы сильнее реагировать на изменяющееся раздражение (появление или исчезновение стимула), тогда как на постоянную стимуляцию она отвечает слабо. При проецировании на сетчатку постоянного изображения с помощью миниатюрного проектора, прикрепленного к контактной линзе, это изображение перестает восприниматься спустя несколько секунд, и вместо него в зрительном ощущении появляется сплошное серое поле. По истечении нескольких секунд восприятие проецируемого на сетчатку изображения восстанавливается, но затем снова исчезает и т. д. 17.3.2. Преобразование энергии света в сетчатке Задняя стенка глаза выстлана эпителием, содержащим черный пигмент меланин, который препятствует отражению света и поглощает его излишки. К пигментному эпителию примыкает слой фоторецепторов, а перед ним расположены слои горизонтальных, биполярных, амакриновых и ганглиозных клеток, чьи аксоны образуют зрительный нерв. В связи с инверсией сетчатки решаются две задачи: 1) обеспечение максимальной прозрачности клеток, расположенных на пути прохождения света к фоторецепторам, 2) выделение в слое фоторецепторов промежутка для прохождения волокон зрительного нерва. Решение первой задачи заключается в анатомическом устройстве центральной ямки, где биполярные и ганглиозные клетки сетчатки располагаются вокруг нее, а их относительно прозрачные безмиелиновые волокна не мешают прохождению света к фоторецепторам. В других областях сетчатки световые лучи отчасти рассеиваются при прохождении сквозь тела клеток, расположенных перед фоторецепторами, но в центральной ямке кванты света без помех попадают на фоторецепторы, представленные здесь преимущественно колбочками. При направленном на определенный объект зрительного поля внимании его изображение всегда фокусируется в центральной ямке для обеспечения максимальной остроты зрения. Сосок зрительного нерва образован аксонами ганглиозных клеток, отсутствие в этой области фоторецепторов делает ее не чувствительной к 17. Сенсорные системы • 723
17.3.2.1. Скотопическая и фотопическая системы сетчатки Фоторецепторный слой сетчатки человека образован примерно 130 миллионами клеток, из которых около семи миллионов являются колбочками, основная масса которых сосредоточена в области центральной ямки, а все остальные фоторецепторы представлены палочками. У обеих разновидностей фоторецепторов существуют три функциональные области: 1) наружный, или внешний, сегмент, ориентированный в направлении эпителиального пигментного слоя и содержащий зрительный пигмент; 2) внутренний сегмент, в котором расположено клеточное ядро и происходят биохимические процессы, связанные с жизнедеятельностью клетки; 3) синаптические окончания, предназначенные для передачи информации от фоторецепторов к биполярным клеткам с помощью медиатора глутамата. Зрительный пигмент палочек родопсин состоит из двух компонентов: это молекула ретиналя, образующаяся из витамина А и способная поглощать свет, а также крупная белковая молекула опсина, не поглощающая свет. Молекула опсина представляет собой извитую цепь из 348 аминокислот, которая семь раз проходит через мембрану зрительного диска, образованного из клеточной мембраны фоторецептора. В наружном сегменте фоторецептора имеется большое количество таких дисков, расположенных подобно стопке поставленных друг на друга монет. Ретиналь существует в темноте как 11-цис-ретиналъ, такая форма изомера идеально соответствует упорядоченному расположению аминокислот в опсине. Энергия поглощенных фотонов превращает ретиналь в 11-транс-изомер, что приводит к конформационным изменениям молекулы опсина и превращению родопсина в нестабильный метародопсин, который сразу же распадается на ретиналь и опсин. Таким образом, действие света уменьшает концентрацию родопсина в фоторецепторе, что приводит к изменениям активности вторичных посредников и величины мембранного потенциала фоторецептора. В темноте происходит ферментативный ресинтез расщепленного родопсина, для которого используется витамин А, поступающий в организм человека с пищей. Родопсин наиболее чувствителен к электромагнитным волнам длиной около 500 нм, но хорошо поглощает и другие волны в диапазоне от 400 до 600 нм. Способность родопсина поглощать волны почти всего светового диапазона позволяет палочкам обеспечить только ахроматическое, т. е. черно-белое, зрение и лишает их возможности различать цвет. Высокое содержание зрительного пигмента в палочках и его способность суммировать фотоны, поглощенные в течение около 100 мс, делает палочки наиболее чувствительными фоторецепторами сетчатки. При слабом сумеречном освещении зрение обеспечивают только палочки, способные возбуждаться вследствие поглощения всего лишь одного фотона. Палочки образуют ско-топическую систему, или систему ночного зрения. Зрительные пигменты колбочек подобны родопсину палочек и состоят
Рис. 17.7. Спектры поглощения четырех разновидностей фоторецепторов. Зрительный пигмент палочек родопсин (Р) имеет максимум поглощения световых волн длиною 496 нм, но способен также к поглощению коротких и длинных волн светового диапазона. Зрительный пигмент колбочек, чувствительных к синему цвету (С), имеет максимум поглощения 419 нм и не поглощает длинные волны оптического диапазона. Пигмент колбочек, чувствительных к зеленому цвету (3), имеет максимум поглощения при 531 нм, а пигмент чувствительных к красному цвету колбочек (К) максимально поглощает волны длиной 596 нм. из светопоглощающей молекулы ретиналя и опсина, который отличается составом аминокислот от белковой части родопсина. Кроме того, колбочки содержат меньшее, чем палочки, количество зрительного пигмента, и для их возбуждения требуется энергия нескольких сотен фотонов. Поэтому колбочки активируются лишь при дневном или достаточно ярком искусственном освещении, они образуют фотопическую систему, или систему дневного зрения. В сетчатке человека существуют три типа колбочек, различающихся между собой по составу аминокислот в опсине зрительного пигмента. Различия в белковой части молекулы определяют особенности взаимодействия каждой из трех форм опсина с ретиналем и специфическую чувствительность к световым волнам разной длины (рис. 17.7). Колбочки одного из трех типов максимально поглощают короткие световые волны с длиной 419 нм, что необходимо для восприятия синего цвета. Другой тип зрительного пигмента наиболее чувствителен к волнам средней длины и имеет максимум поглощения при 531 нм, он служит для восприятия зеленого цвета. Третий тип зрительного пигмента максимально поглощает длинные волны с максимумом при 559 нм, что позволяет воспринимать красный цвет. Наличие трех типов колбочек обеспечивает человеку восприятие всей цветовой палитры, в которой существует свыше семи миллионов цветовых градаций, тогда как скотопическая система палочек позволяет различать лишь около пятисот черно-белых градаций. 17. Сенсорные системы • 725
Фоторецепторы образуют синапсы с биполярными клетками двух типов, различающихся по способу управления хемозависи-мыми натриевыми каналами в синапсах. Действие глутамата приводит к открытию каналов для ионов натрия и деполяризации мембраны одних биполярных клеток и к закрытию натриевых каналов и гиперполяризации биполярных клеток другого типа. Наличие двух типов биполярных клеток необходимо для формирования антагонизма между центром и периферией рецептивных полей ганглиоз-ных клеток.
17.3.2.3. Адаптация Временное ослепление при быстром переходе от темноты к яркому освещению исчезает спустя несколько секунд благодаря процессу световой адаптации. Одним из механизмов световой адаптации является рефлекторное сужение 726 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Способность различать окружающие предметы исчезает на некоторое время при быстром переходе от яркого света к темноте. Она постепенно восстанавливается в ходе темповой адаптации, обусловленной расширением зрачков и переключением зрительного восприятия с фотопической системы на скотопическую. Темновую адаптацию палочек определяют медленные изменения функциональной активности белков, приводящие к повышению их чувствительности. В механизме темновой адаптации участвуют и горизонтальные клетки, способствующие увеличению центральной части рецептивных полей в условиях низкой освещенности.
|
|||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-21; просмотров: 1443; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.214 (0.013 с.) |


 Роговица Точка фиксации
Роговица Точка фиксации близких предметов и в обоих случаях фокусирует их изображение на сетчатку, называется аккомодацией. Аккомодацию обеспечивают изменения кривизны хрусталика, зависимой от степени натяжения его тонкой и прозрачной капсулы, которая переходит по краям в циннову связку, прикрепляющуюся к ресничному телу. Гладкие мышцы ресничного тела, управляемые парасимпатическими нейронами, регулируют натяжение цинновой связки: при полном расслаблении мышц связка натягивает капсулу хрусталика, заставляя его принимать максимально уплощенную форму, необходимую для рассмотрения далеких предметов. При сокращении ресничных мышц натяжение цинновой связки уменьшается, хрусталик в силу своей эластичности принимает более выпуклую форму и поэтому сильнее преломляет световые лучи, что, например, происходит во время чтения этого текста.
близких предметов и в обоих случаях фокусирует их изображение на сетчатку, называется аккомодацией. Аккомодацию обеспечивают изменения кривизны хрусталика, зависимой от степени натяжения его тонкой и прозрачной капсулы, которая переходит по краям в циннову связку, прикрепляющуюся к ресничному телу. Гладкие мышцы ресничного тела, управляемые парасимпатическими нейронами, регулируют натяжение цинновой связки: при полном расслаблении мышц связка натягивает капсулу хрусталика, заставляя его принимать максимально уплощенную форму, необходимую для рассмотрения далеких предметов. При сокращении ресничных мышц натяжение цинновой связки уменьшается, хрусталик в силу своей эластичности принимает более выпуклую форму и поэтому сильнее преломляет световые лучи, что, например, происходит во время чтения этого текста. этой аномалии рефракции необходимы очки с двояковыпуклыми линзами, преломляющими световые лучи так, чтобы они фокусировались на сетчатке.
этой аномалии рефракции необходимы очки с двояковыпуклыми линзами, преломляющими световые лучи так, чтобы они фокусировались на сетчатке.



 Первичная зрительная кора
Первичная зрительная кора 17.3.1.5. Движения глаз
17.3.1.5. Движения глаз свету, в связи с чем она получила название слепого пятна. На соски правого и левого глаза благодаря диспарации попадают неодинаковые части проецируемого изображения, поэтому в условиях естественного зрительного восприятия пробел изображения на сетчатке не осознается. Кроме того, быстрые движения глаз позволяют получать нужную информацию о любой области рассматриваемого изображения, и отсутствие сигналов от слепого пятна замещается информацией, поступившей от ближайших к нему фоторецепторов.
свету, в связи с чем она получила название слепого пятна. На соски правого и левого глаза благодаря диспарации попадают неодинаковые части проецируемого изображения, поэтому в условиях естественного зрительного восприятия пробел изображения на сетчатке не осознается. Кроме того, быстрые движения глаз позволяют получать нужную информацию о любой области рассматриваемого изображения, и отсутствие сигналов от слепого пятна замещается информацией, поступившей от ближайших к нему фоторецепторов.
 17.3.2.2. Рецепторный потенциал палочек и колбочек
17.3.2.2. Рецепторный потенциал палочек и колбочек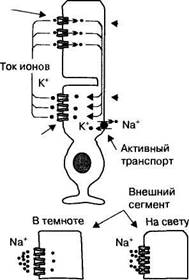 Специфической особенностью фоторецепторов является темновой ток катионов через открытые мембранные каналы внешних сегментов (рис. 17.8). Эти каналы открываются при высокой концентрации циклического гуано-зинмонофосфата, который является вторичным посредником рецепторного белка (зрительного пигмента). Темновой ток катионов деполяризует мембрану фоторецептора до приблизительно —40 мВ, что приводит к выделению медиатора в его синаптическом окончании. Активированные поглощением света молекулы зрительного пигмента стимулируют активность фос-фодиэстеразы — фермента, расщепляющего цГМФ, поэтому при действии света на фоторецепторы в них уменьшается концентрация цГМФ. В результате управляемые этим посредником катионные каналы закрываются, и ток катионов в клетку прекращается. Вследствие непрерывного выхода ионов калия из клеток, мембрана фоторецепторов гиперполяризуется приблизительно до —70 мВ, эта гиперполяризация мембраны является рецепторным потенциалом. При возникновении рецепторного потенциала прекращается выделение глутамата в синаптических окончаниях фоторецептора.
Специфической особенностью фоторецепторов является темновой ток катионов через открытые мембранные каналы внешних сегментов (рис. 17.8). Эти каналы открываются при высокой концентрации циклического гуано-зинмонофосфата, который является вторичным посредником рецепторного белка (зрительного пигмента). Темновой ток катионов деполяризует мембрану фоторецептора до приблизительно —40 мВ, что приводит к выделению медиатора в его синаптическом окончании. Активированные поглощением света молекулы зрительного пигмента стимулируют активность фос-фодиэстеразы — фермента, расщепляющего цГМФ, поэтому при действии света на фоторецепторы в них уменьшается концентрация цГМФ. В результате управляемые этим посредником катионные каналы закрываются, и ток катионов в клетку прекращается. Вследствие непрерывного выхода ионов калия из клеток, мембрана фоторецепторов гиперполяризуется приблизительно до —70 мВ, эта гиперполяризация мембраны является рецепторным потенциалом. При возникновении рецепторного потенциала прекращается выделение глутамата в синаптических окончаниях фоторецептора. зрачков, другой зависит от концентрации ионов кальция в колбочках. При поглощении света в мембранах фоторецепторов закрываются катионные каналы, что прекращает вхождение ионов натрия и кальция и уменьшает их внутриклеточную концентрацию. Высокая концентрация ионов кальция в темноте подавляет активность гуанилатциклазы — фермента, определяющего образование цГМФ из гуанозинтрифосфата. Вследствие снижения концентрации кальция, обусловленного поглощением света, активность гуанилатциклазы повышается, что ведет к дополнительному синтезу цГМФ. Повышение концентрации этого вещества приводит к открытию катионных каналов, восстановлению тока катионов в клетку и, соответственно, способности колбочек отвечать на световые раздражители как обычно. Низкая концентрация ионов кальция способствует десенситизации колбочек, т. е. уменьшению их чувствительности к свету. Десенситизация обусловлена изменением свойств фосфодиэстеразы и белков катионных каналов, становящихся менее чувствительными к концентрации цГМФ.
зрачков, другой зависит от концентрации ионов кальция в колбочках. При поглощении света в мембранах фоторецепторов закрываются катионные каналы, что прекращает вхождение ионов натрия и кальция и уменьшает их внутриклеточную концентрацию. Высокая концентрация ионов кальция в темноте подавляет активность гуанилатциклазы — фермента, определяющего образование цГМФ из гуанозинтрифосфата. Вследствие снижения концентрации кальция, обусловленного поглощением света, активность гуанилатциклазы повышается, что ведет к дополнительному синтезу цГМФ. Повышение концентрации этого вещества приводит к открытию катионных каналов, восстановлению тока катионов в клетку и, соответственно, способности колбочек отвечать на световые раздражители как обычно. Низкая концентрация ионов кальция способствует десенситизации колбочек, т. е. уменьшению их чувствительности к свету. Десенситизация обусловлена изменением свойств фосфодиэстеразы и белков катионных каналов, становящихся менее чувствительными к концентрации цГМФ.


