
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Переработка сенсорной информации в слуховой кореСодержание книги
Поиск на нашем сайте
В первичной слуховой коре (поле 41) кортикальные колонки расположены тонотопически для раздельной переработки информации о звуках различной частоты слухового диапазона. Они также содержат нейроны, которые избирательно реагируют на звуки различной продолжительности, на повторяющиеся звуки, на шумы с широким частотным диапазоном и т. п. В слуховой коре происходит объединение информации о высоте тона и его интенсивности, о временных интервалах между отдельными звуками. Вслед за этапом регистрации и объединения элементарных признаков звукового раздражителя, который осуществляют простые нейроны, в переработку информации включаются комплексные нейроны, избирательно реагирующие только на узкий диапазон частотных или амплитудных модуляций звука. Подобная специализация нейронов позволяет слуховой системе создавать целостные слуховые образы, с характерными только для них сочетаниями элементарных компонентов слухового раздражителя. Такие со- 17. Сенсорные системы • 743
17.5. Вестибулярная сенсорная система Функция вестибулярной сенсорной системы состоит в обеспечении мозга информацией о положении головы в пространстве, о действии гравитации и сил, вызывающих линейные или угловые ускорения. Эта функция необходима для поддержания равновесия, т. е. устойчивого положения тела в пространстве, и для пространственной ориентации человека. Вестибулярная система включает в себя периферический отдел, состоящий из расположенного во внутреннем ухе вестибулярного аппарата, проводящие пути, переключательные центры, представленные вестибулярными ядрами продолговатого мозга и таламусом, и проекционную область коры в постцентральной извилине. Адекватными раздражителями вестибулярной системы являются гравитация и силы, сообщающие телу линейное или угловое ускорение. Специфическая особенность вестибулярной системы состоит в том, что значительная часть перерабатываемой в ней сенсорной информации используется для автоматической регуляции функций, осуществляемой без сознательного контроля. Вестибулярная система взаимодействует на нескольких уровнях своей иерархической организации со зрительной и со-матосенсорной системами; три эти системы дополняют друг друга в предоставлении человеку информации, необходимой для его пространственной ориентации.
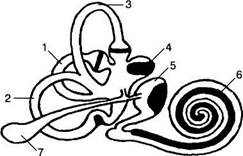
17.5.1. Вестибулярный аппарат Внутри костных лабиринтов, расположенных в пирамидах височных костей, имеются прикрепленные к ним соединительнотканными тяжами перепончатые лабиринты, заполненные вязкой жидкостью эндолимфы (рис. 17.15). Лабиринт образован двумя отолитовыми органами и тремя полукружными каналами, расположенными в трех плоскостях: горизонтальный канал, передний вертикальный канал — во фронтальной плоскости и задний вертикальный канал — в сагиттальной плоскости. Все три канала соединены в полости преддверия, от латинского определения которого (уе$11Ъи1ит) происходит само название вестибулярного аппарата. В месте соединения с преддверием каналы расширены в виде ампул, в которых содержится ре-цепторный эпителий, выступающий внутрь в форме гребня или кристы. Каждая криста покрыта купулой, представляющей собой аморфное желеобразное вещество, скрепленное множеством фибриллярных волокон. При угловых ускорениях, когда в силу инерции происходит сдвиг эндолимфы, купула тоже смещается, что приводит к деформации погруженных в нее вторичных рецепторных клеток с последующим возникновением в них ре-цепторного потенциала. В полости преддверия имеются два расширения: мешочек ($асси1и$) и маточка (шпси1и$), сообщающиеся между собой с помощью У-образного протока и представляющие собой отолитовые органы. Рецепторный эпителий маточки и мешочка расположен на небольших возвышениях — маку-лах, покрытых отолитовой мембраной, которая имеет слоистое строение и содержит множество мелких, но тяжелых кристаллов карбоната кальция {отолиты или отокинии). Макула маточки расположена в горизонтальной 744 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Т Т!Т Т ИмпульсацИя в афферентных Окончаниях вестибулярного нерва
| Возбуждение Торможение Рис. 17.15. Вестибулярная сенсорная система. А. Схема перепончатого лабиринта, рецепторные области выделены черным цветом: 1) горизонтальный канал; 2) задний вертикальный канал; 3) передний вертикальный канал; 4) маточка; 5) мешочек; 6) канал улитки; 7) эндолимфатический мешочек. Б. Волосковые рецепторные клетки вестибулярного аппарата: показана зависимость между направлением смещения волосков и активностью клеток. плоскости (при вертикальном положении головы), а макула мешочка ориентирована вертикально. При действии силы тяжести или линейного ускорения отолитовые мембраны сдвигаются относительно макул, а вследствие этого раздражаются имеющиеся в них вторичные рецепторные клетки. Таким образом, устройство вестибулярного аппарата обеспечивает возбуждение вторичных рецепторных клеток благодаря действию силы тяжести и прямолинейного ускорения (макулы мешочка и маточки) и вследствие угловых ускорений (купулы полукружных каналов). 17.5.1.1. Свойства рецепторных клеток вестибулярного аппарата Рецепторы макул и купул представлены волосковыми клетками, являющимися вторичными механорецепторами и образующими синапсы с периферическими окончаниями нейронов вестибулярного ганглия (первичные сенсорные нейроны). Каждый рецептор имеет пучок из 40—80 волосков — стереоцилий, достигающих в длину 50 мкм, а также расположенный экс- 17. Сенсорные системы • 745
Нейроны вестибулярного ганглия, образующие синапсы на основании рецепторов, обладают спонтанной фоновой активностью, характер которой меняется под влиянием медиаторов волосковых клеток, которыми предположительно могут быть глутамат и/или ГАМК. Рецептивные поля нейронов вестибулярного ганглия включают в среднем три волосковых клетки ампул полукружных каналов либо 4—6 рецепторов макул маточки или мешочка. 17.5.1.2. Адекватные раздражители рецепторов отолитовых органов Рецепторы маточки и мешочка служат датчиками гравитации и линейных ускорений. При вертикальном положении головы человека макула маточки расположена в горизонтальной плоскости, при наклоне головы покрывающая макулу отолитовая мембрана смещается, подчиняясь силе тяжести. Смещение отолитовой мембраны сгибает стереоцилий рецепторных клеток, отвечающих на деформацию образованием рецепторного потенциала. В зависимости от направления и степени наклона головы сильнее других возбуждаются такие рецепторы, функциональная поляризация которых соответствует именно этому направлению, прочие рецепторы возбуждаются слабее или же тормозятся. Изменившееся соотношение возбужденных и заторможенных рецепторных клеток вызывает в зависимых от них нейронах вестибулярного ганглия адекватное изменение фоновой активности. Нейроны вестибулярного ганглия передают полученную от рецепторных клеток информацию в центральную нервную систему. Указанные процессы возникают не только при наклоне головы, но и при любом отклонении положения всего тела от вертикальной оси, например при спортивной или профессиональной деятельности, случайном падении, использовании аттракционов (качели, американские горки).
Макула мешочка при вертикальном положении тела и головы расположена в вертикальной плоскости, и ее отолитовая мембрана сдвигается при действии линейных ускорений, вызывая раздражение рецепторов. В зависимости от направления, в котором происходит линейное ускорение, возбуждаются наиболее чувствительные именно к нему рецепторы. Наличие нескольких популяций рецепторов, различающихся своей функциональной поляризацией, позволяет им в целом передавать сенсорным нейронам информацию о линейных перемещениях в любом направлении. Чувствительность этих рецепторов позволяет человеку ощутить прямолинейное ускорение, превысившее 2 см/с2 и наклон головы в сторону всего на Г. Наряду с этим рецепторный аппарат мешочка высокочувствителен к действию вибрации. 746 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Угловые ускорения возникают при вращении тела вокруг одной из трех пространственных осей, расположенных перпендикулярно друг другу, они возникают также при поворотах головы и ее наклонах. При вращении вокруг вертикальной оси кресла с сидящим человеком у него раздражаются рецепторы горизонтального канала. Раздражение возникает в самом начале вращения, когда инертная эндолимфа остается неподвижной, что создает усилие, смещающее купулу в противоположную вращению сторону. При остановке вращения движение эндолимфы по инерции продолжается, что вызывает смещение купулы в сторону уже прекратившегося движения. В результате смещения купулы стереоцилии горизонтального канала сгибаются сначала в одном направлении, что сопровождается деполяризацией волосковых клеток, а затем — в противоположном направлении, что вызывает гиперполяризацию рецепторов. Соответственно этому волосковая клетка увеличивает или уменьшает выделение медиатора, действующего на окончание нейрона вестибулярного ганглия, что повышает или понижает его фоновую активность.
При вращении вокруг осей, перпендикулярных фронтальной или сагиттальной плоскостям, аналогичные вышеописанным изменения активности рецепторов происходят в переднем или заднем вертикальных полукружных каналах. Вращение вокруг какой-либо диагональной оси вызывает движение эндолимфы в двух каналах одновременно, соответственно реагируют рецепторы, расположенные в купулах обоих каналов. Наличие трех полукружных каналов обеспечивает человеку восприятие вращения и поворотов головы в любой плоскости трехмерного пространства. Сенсорная чувствительность вестибулярной системы позволяет ощущать угловые ускорения, достигшие значения 2°/с2. 17.5.2. Центральная часть вестибулярной системы Центральные аксоны первичных сенсорных нейронов вестибулярного ганглия оканчиваются на нейронах вестибулярных ядер: верхнего (ядро Бехтерева), нижнего (ядро Роллера), латерального (ядро Дейтерса) и медиального (ядро Швальбе). Эти ядра представляют собой единый функциональный комплекс, в котором объединяется афферентная информация от вестибулярных ганглиев и от проприоцепторов; эта афферентация определяет характер активности нейронов вестибулярных ядер. Вестибулярные ядра являются подкорковыми центрами познотонических и статокинетиче-ских рефлексов (регуляция равновесия), глазодвигательных рефлексов (зрительное наблюдение окружающего мира) и опосредованных через гипоталамус вестибуловисцеральных реакций, с механизмом которых связаны проявления кинетозов — морской болезни, проявляющейся головокружением и тошнотой. Осознаваемое восприятие изменений положения головы происходит в результате последовательной переработки информации сначала в вестибулярных ядрах, затем в задних вентральных ядрах таламуса, образующих проекцию к постцентральным извилинам. Дополнительная информация поступает в проекционную кору непрямым путем: от вестибулярных ядер в мозжечок, а из него к вентролатеральным ядрам таламуса и проекционной коре. Первичная проекционная область вестибулярной чувствительности размещена в задней центральной извилине преимущественно той стороны тела, на которой расположен вестибулярный аппарат. Еще одна проекция, 17. Сенсорные системы • 747
17.6. Вкусовая сенсорная система
17.6.1. Вкусовая рецепция
Выпячивания слизистой оболочки языка (сосочки) содержат у человека около 2000 вкусовых почек, каждая из которых образована 30—60 клетками, расположенными в почке наподобие долек апельсина. Рецепторные клетки составляют около 5—7 % всех клеток вкусовой почки, они от- Лицевой нерв
Рис. 17.16. Рецепторы и афферентные пути вкусовой системы.
Рецепторные клетки вкусовой почки имеют ворсинки, выступающие во вкусовую пору. Они реагируют на присоединение химических веществ изменением количества медиатора, действующего на афферентные волокна нейронов первого порядка. Афферентные сигналы поступают в ЦНС ипсилатерально по волокнам лицевого, языкоглоточного и блуждающего нервов к нейронам второго порядка, расположенным в ядре одиночного пучка. 748 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
17.6.1.1. Рецепторные потенциалы вкусовых клеток Мембрана микроворсинок вкусовых клеток содержит специфические участки (рецепторы), предназначенные для связывания растворенных в жидкой среде полости рта химических молекул. Существует четыре разновидности вкусовых ощущений, или четыре вкусовые модальности: сладкое, кислое, соленое и горькое. Строгой зависимости между химической природой вещества и вкусовым ощущением нет: например, сладким вкусом обладают не только сахара, но и некоторые неорганические соединения (соли свинца, бериллия), а самым сладким веществом является не усваиваемый организмом сахарин. Большинство вкусовых клеток полимодальны, т. е. могут реагировать на стимулы всех четырех вкусовых модальностей. Присоединение к специфическим рецепторам молекул, обладающих сладким вкусом, активирует систему вторичных посредников аденилатцик-лазы — циклического аденозинмонофосфата, которые закрывают мембранные каналы ионов калия, и поэтому мембрана рецепторной клетки деполяризуется. Вещества, обладающие горьким вкусом, активируют одну из двух систем вторичных посредников: 1) фосфолипазу С — инозитол-3-фос-фат, что приводит к выходу из внутриклеточного депо ионов кальция с последующим выделением медиатора из рецепторной клетки; 2) специфический С-белок гастдуцин, регулирующий внутриклеточную концентрацию цАМФ, которая управляет катионными каналами мембраны и этим определяет возникновение рецепторного потенциала. Действие на рецепторы молекул, имеющих соленый вкус, сопровождается открытием управляемых натриевых каналов и деполяризацией вкусовой клетки. Вещества, обладающие кислым вкусом, закрывают мембранные каналы для ионов калия, что ведет к деполяризации рецепторной клетки. Величина рецепторного потенциала зависит от вкусового качества и концентрации химического вещества, действующего на клетку. Возникновение рецепторного потенциала приводит к выделению вкусовой клеткой медиатора, действующего через синапс на афферентное волокно первичного сенсорного нейрона, в котором через 40—50 мс от начала действия стимула повышается частота потенциалов действия. Возникшие в афферентных волокнах нервные импульсы проводятся к ядрам одиночных пучков продолговатого мозга. При повышении концентрации действующего вещества общее количество реагирующих чувствительных волокон возрастает за счет вовлечения высокопороговых афферентов в передачу информации от рецепторов. 17.6.1.2. Вкусовая чувствительность Пороги вкусовой чувствительности выявляются поочередным нанесением на поверхность языка растворов веществ, обладающих разными вкусовыми качествами (табл. 17.4). Абсолютным порогом чувствительности считают появление определенного вкусового ощущения, отличающегося от вкуса 17. Сенсорные системы • 749
дистиллированной воды. Вкус одного и того же вещества может восприниматься по-разному в зависимости от его концентрации в растворе; например, при малой концентрации хлорида натрия он ощущается сладким, а при большей концентрации — соленым. Максимальная способность различать концентрацию растворов одного и того же вещества и, соответственно, самый низкий дифференциальный порог вкусовой чувствительности характерны для среднего диапазона концентраций, а при высоких концентрациях вещества дифференциальный порог повышается. Абсолютные пороги вкусовой чувствительности индивидуально различаются, но у подавляющего большинства людей самым низким оказывается порог определения веществ с горьким вкусом. Эта особенность восприятия возникла в эволюции, она способствует отказу от употребления в пищу веществ горького вкуса, к которым принадлежат алкалоиды многих ядовитых растений. Вкусовые пороги различаются у одного и того же человека в зависимости от его потребности в тех или иных веществах, они повышаются вследствие длительного употребления веществ с характерным вкусом (например, сладостей или соленостей) или курения, потребления алкоголя, обжигающих напитков. Разные области языка различаются вкусовой чувствительностью к различным веществам, что обусловлено особенностями распределения вкусовых рецепторов. Кончик языка более других областей чувствителен к сладкому, боковые стороны языка — к кислому и соленому, а корень языка — к горькому. Вкусовые ощущения в большинстве случаев мультимодальны и основаны не только на избирательной химической чувствительности вкусовых рецепторных клеток, но и на раздражении пищей терморецепторов и механорецепторов ротовой полости, а также действии летучих компонентов пищи на обонятельные рецепторы. 17.6.2. Центральный отдел вкусовой системы Афферентную иннервацию вкусовых почек передних 2/з языка осуществляют чувствительные волокна лицевых нервов (барабанная струна), вкусовые рецепторы задней трети языка и твердого неба образуют синапсы с чувствительными волокнами языкоглоточных нервов, а передачу сигналов от небольшого количества вкусовых рецепторов в глотке и надгортаннике осуществляют чувствительные волокна блуждающих нервов. Волокна барабанной струны полимодальны и реагируют повышением частоты нервных импульсов на действие веществ с любым вкусовым качеством. Однако если одни волокна максимально возбуждаются при действии веществ со сладким вкусом, то наибольшая активность других волокон возникает в ответ на действие веществ соленого вкуса, либо кислого или горького вкуса. Если определить активность одиночного волокна к веществам каждой из четырех вкусовых субмодальностей, то можно получить «вкусовой профиль» такого волокна, который будет отличаться от вкусовых профилей других волокон. 750 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Электрическая активность нейронов ядра одиночного пучка повышается в ответ на действие вкусовых раздражителей разных модальностей, а также вследствие тактильного и температурного раздражения языка и ротовой полости. Эти нейроны так же, как и нейроны первого порядка, мультимодальны, однако каждый такой нейрон отличается своей индивидуальной активностью. Различия состоят в том, что максимальная или минимальная активность нейрона проявляется в ответ на действие веществ, обладающих разным вкусом, что формирует индивидуальный профиль вкусовой активности данного нейрона. Указанные индивидуальные различия сохраняются у нейронов вентрального ядра таламуса, и лишь в коре существуют клетки, реагирующие только на действие вещества с определенным вкусовым качеством, т. е. обладающие специфической вкусовой чувствительностью. Наряду с ними в проекционной коре имеются нейроны, реагирующие не только на действие вкусовых раздражителей, но и на тактильное и температурное раздражение языка, что способствует целостному восприятию всех качеств пищи. Вкусовое восприятие Вкусовые ощущения отчетливо проявляются уже у новорожденных детей, а грудной ребенок 4—5 мес хорошо различает вкусовые изменения пищи. Вкусовое восприятие человека формируется в результате соединения муль-тимодальных сенсорных ощущений и механизмов памяти, хранящей индивидуальный опыт и связанной с усвоенными традициями питания. Изменения вкусового восприятия часто связаны с нарушениями обоняния, а не собственно вкусовой чувствительности. Ротовая полость сообщается с носовой, и пахучие вещества пищи достигают обонятельных рецепторов, поэтому представление о вкусе пищи почти всегда основано не только на вкусовом, но и на обонятельном восприятии. Гастрономические предпочтения людей связаны с комплексным действием тех или иных блюд, включающем не только вкусовые и обонятельные, но также тактильные температурные и зрительные ощущения.
|
||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-21; просмотров: 411; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.118.9.147 (0.013 с.) |

 четания могут быть зафиксированы энграммами памяти, что в дальнейшем позволяет сравнивать новые акустические стимулы с прежними. Некоторые комплексные нейроны слуховой коры возбуждаются сильнее всего в ответ на действие звуков человеческой речи.
четания могут быть зафиксированы энграммами памяти, что в дальнейшем позволяет сравнивать новые акустические стимулы с прежними. Некоторые комплексные нейроны слуховой коры возбуждаются сильнее всего в ответ на действие звуков человеческой речи.
 Смещение волосков
Смещение волосков В покое
В покое центрично по отношению к стереоцилиям один длинный волосок — кино-цилию. Если пучок стереоцилий наклоняется под влиянием механического стимула в сторону киноцилии, рецептор деполяризуется, а при отклонении стереоцилий от киноцилии происходит гиперполяризация мембраны рецептора. Вследствие этого при сгибании пучка стереоцилий в одном направлении волосковая клетка возбуждается, а при сгибании этого же пучка в противоположном направлении — тормозится, т. е. у каждой волосковой клетки выявляются два функциональных полюса. Направление функциональной поляризации изменяется от одной клетки к другой, а рецептор-ный эпителий в целом содержит полный комплект клеток для регистрации стимулов, действующих в любом возможном направлении.
центрично по отношению к стереоцилиям один длинный волосок — кино-цилию. Если пучок стереоцилий наклоняется под влиянием механического стимула в сторону киноцилии, рецептор деполяризуется, а при отклонении стереоцилий от киноцилии происходит гиперполяризация мембраны рецептора. Вследствие этого при сгибании пучка стереоцилий в одном направлении волосковая клетка возбуждается, а при сгибании этого же пучка в противоположном направлении — тормозится, т. е. у каждой волосковой клетки выявляются два функциональных полюса. Направление функциональной поляризации изменяется от одной клетки к другой, а рецептор-ный эпителий в целом содержит полный комплект клеток для регистрации стимулов, действующих в любом возможном направлении. 17.5.1.3. Адекватные раздражители рецепторов полукружных каналов
17.5.1.3. Адекватные раздражители рецепторов полукружных каналов отличающаяся двусторонним представительством вестибулярной чувствительности, имеется во вторичной моторной коре. Осознание пространственного расположения и схемы тела происходит с участием заднетеменных регионов коры (поля 5 и 7), где осуществляется интеграция вестибулярной, зрительной и соматосенсорнои чувствительности человека.
отличающаяся двусторонним представительством вестибулярной чувствительности, имеется во вторичной моторной коре. Осознание пространственного расположения и схемы тела происходит с участием заднетеменных регионов коры (поля 5 и 7), где осуществляется интеграция вестибулярной, зрительной и соматосенсорнои чувствительности человека.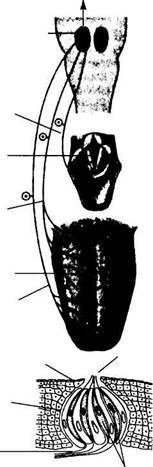 Функция вкусовой сенсорной системы состоит в оценке качества пищи или отвергаемых веществ. Адекватным раздражителем вкусовой системы служат молекулы органических или неорганических веществ, которые поступают в полость рта при приеме пищи и присоединяются к хеморецеп-торным клеткам языка и ротовой полости. Эти клетки являются вторичными рецепторами и обладают высокой химической избирательностью, а присоединение химических веществ вызывает в них образование рецептор-ного потенциала, что сопровождается выделением медиатора, действующего на чувствительные окончания первичных сенсорных нейронов. В передаче сигналов от рецепторов в центральную нервную систему участвуют афферентные волокна IX, VII и X черепных нервов (рис. 17.16). Центральный отдел вкусовой сенсорной системы образуют ядра одиночных пучков продолговатого мозга и вентральные ядра таламуса, образующие проекцию в первичную проекционную кору. Проекционная вкусовая кора соответствует области представительства языка в постцентральных извилинах.
Функция вкусовой сенсорной системы состоит в оценке качества пищи или отвергаемых веществ. Адекватным раздражителем вкусовой системы служат молекулы органических или неорганических веществ, которые поступают в полость рта при приеме пищи и присоединяются к хеморецеп-торным клеткам языка и ротовой полости. Эти клетки являются вторичными рецепторами и обладают высокой химической избирательностью, а присоединение химических веществ вызывает в них образование рецептор-ного потенциала, что сопровождается выделением медиатора, действующего на чувствительные окончания первичных сенсорных нейронов. В передаче сигналов от рецепторов в центральную нервную систему участвуют афферентные волокна IX, VII и X черепных нервов (рис. 17.16). Центральный отдел вкусовой сенсорной системы образуют ядра одиночных пучков продолговатого мозга и вентральные ядра таламуса, образующие проекцию в первичную проекционную кору. Проекционная вкусовая кора соответствует области представительства языка в постцентральных извилинах. личаются от остальных клеток (опорных, базалъных) наличием микроворсинок, выступающих во вкусовую пору — отверстие на вершине вкусовой почки. Продолжительность жизни рецепторной клетки составляет 10—12 дней, разрушенные клетки заменяются новыми, образующимися из делящихся базальных клеток и устанавливающими точно такие же, как у своей предшественницы, синаптические контакты с 1—6 отростками первичного сенсорного нейрона, поэтому специфичность вкусовой чувствительности рецепторов после регенерации не изменяется.
личаются от остальных клеток (опорных, базалъных) наличием микроворсинок, выступающих во вкусовую пору — отверстие на вершине вкусовой почки. Продолжительность жизни рецепторной клетки составляет 10—12 дней, разрушенные клетки заменяются новыми, образующимися из делящихся базальных клеток и устанавливающими точно такие же, как у своей предшественницы, синаптические контакты с 1—6 отростками первичного сенсорного нейрона, поэтому специфичность вкусовой чувствительности рецепторов после регенерации не изменяется. Сенсорные нейроны второго порядка содержатся в составе ядра одиночного пучка, вытянутого вдоль продолговатого мозга. На этих нейронах оканчиваются чувствительные волокна лицевого, языкоглоточного и блуждающего нервов, а их аксоны переходят на противоположную сторону в составе медиального лемниска и оканчиваются на нейронах вентрального ядра таламуса. Нейроны таламуса образуют проекцию на латеральную часть постцентральной извилины — корковую проекционную область вкусовой чувствительности.
Сенсорные нейроны второго порядка содержатся в составе ядра одиночного пучка, вытянутого вдоль продолговатого мозга. На этих нейронах оканчиваются чувствительные волокна лицевого, языкоглоточного и блуждающего нервов, а их аксоны переходят на противоположную сторону в составе медиального лемниска и оканчиваются на нейронах вентрального ядра таламуса. Нейроны таламуса образуют проекцию на латеральную часть постцентральной извилины — корковую проекционную область вкусовой чувствительности.


