
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Переработка информации в переключательных ядрах и проводящих путях сенсорной системыСодержание книги
Поиск на нашем сайте
Соматовисцеральная сенсорная система Соматовисцеральная система перерабатывает сенсорную информацию нескольких модальностей, получая ее от кожи, мышц, суставов, внутренних органов и кровеносных сосудов. Она содержит разные типы рецепторов: механорецепторы, терморецепторы, хеморецепторы, осморецепторы, но-цицепторы. Поступающая информация передается по афферентным волокнам в составе чувствительных и смешанных нервов, перерабатывается раздельными проводящими путями и переключательными ядрами, а затем доставляется в соматосенсорную кору, занимающую постцентральные извилины мозга и организованную топически. Тактильная чувствительность Часть соматовисцеральной системы, обеспечивающая чувство осязания, включает несколько разновидностей механорецепторов кожи, представленных свободными нервными окончаниями либо инкапсулированными, т. е. заключенными в капсулу из соединительной ткани или видоизмененных клеток эпидермиса (рис. 17.4). Свободные нервные окончания иннервиру-ют волосяные фолликулы пушковых волос, покрывающих большую часть тела человека, а также грубых волос, растущих на голове, в подмышечных впадинах, на лобке, а у мужчин еще и на лице. Свободные нервные окон- 712 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Среди инкапсулированных окончаний различают тельца Пачини, Мейс-снера, Руффини, диски Меркеля, тактильные тельца Пинкуса — Иon-о, колбы Краузе. В зависимости от строения и формы капсулы нервные окончания подвержены наиболее сильному воздействию либо в результате давления действующим перпендикулярно раздражителем, либо вследствие бокового смещения капсулы, которая играет роль механического преобразователя энергии внешних стимулов. Большинство инкапсулированных рецепторов содержится в лишенной волос коже пальцев рук и ног, ладоней и подошв, лица, губ, языка, сосков и половых органов, где они распределены с различной плотностью и на разной глубине. Тельца Пачини имеются также в сухожилиях, связках и брыжейке. Механорецепторы кожи различаются по скорости адаптации к действующему раздражителю. Быстроадаптирующиеся (фазные) рецепторы возбуждаются только в момент смещения кожи и волос и служат датчиками скорости воздействия стимула. Это свойство присуще тельцам Мейснера, рецепторам волосяных фолликулов и особенно тельцам Пачини, способным реагировать на изменения скорости продолжающего свое действие стимула. Медленно адаптирующиеся (тонические) рецепторы не прекращают генерировать потенциалы действия при продолжительном действии 17. Сенсорные системы • 713
раздражителя, если он оказывает давление Таблица 17.1. Пространственна кожу: такие рецепторы служат датчиками НЬ|Й двухточечный порог в раз-интенсивности действующего стимула ных У430™8"тела (тельца Руффини, диски Меркеля). Площадь рецептивных полей сенсорных нейронов, иннервирующих тельца Мейсне-ра и диски Меркеля, составляет в среднем около 12 мм2, а у нейронов с окончаниями в виде телец Пачини и Руффини она на порядок больше. Рецептивные поля различающихся своими рецепторами сенсорных нейронов перекрываются, поэтому при действии на кожу комплекса стимулов одновременно возбуждаются разные виды рецепторов, что позволяет ощущать все динамические и статические свойства такого комплекса. Обработка и анализ информации сигналов от различных рецепторов происходит на высших уровнях сенсорной системы, формирующих комплексное восприятие действующих на поверхность тела стимулов. Плотность меха-норецепторов в разных участках кожи не одинакова, чем определяются разные показатели пространственного дифференциального порога, т. е. наименьшего расстояния между двумя точками, раздражение каждой из которых ощущается раздельно (табл. 17.1). Приведенные в таблице данные не следует считать эталоном, поскольку дифференциальная чувствительность различается у разных людей. Инкапсулированные рецепторы иннервируются миелинизированными волокнами первичных сенсорных нейронов, которые проводят нервные импульсы в ЦНС со скоростью около 30—70 м/с. Немиелинизированные волокна передают потенциалы действия от свободных нервных окончаний со значительно меньшей скоростью — около 1 м/с, поэтому ощущение действующего на них стимула возникает относительно позже. Центральные отростки первичных сенсорных нейронов входят в спинной мозг в составе задних корешков и разделяются в задних рогах спинного мозга на коллатерали. Восходящие коллатерали достигают переключательных ядер заднего столба продолговатого мозга, откуда специфическая информация передается на противоположную сторону мозга по лемнисковому пути, поступает к проекционным ядрам таламуса, а затем в соматосенсорную кору, с участием которой формируется чувство осязания. 17.2.2. Проприоцептивная чувствительность Проприоцептивные ощущения дают человеку возможность воспринимать изменения положения отдельных частей тела в покое и во время совершаемых движений. Информация, поступающая от проприоцепторов, позволяет ему постоянно контролировать позу и точность произвольных движений, дозировать силу мышечных сокращений при противодействии внешнему сопротивлению, например при подъеме или перемещении груза. Проприоцепторы представлены интрафузалъными волокнами мышц (датчики длины мышцы), сухожильными рецепторами Гольджи (датчики напряжения мышцы) и механорецепторами суставных капсул, отвечающими изменениями фоновой активности на сгибание или разгибание в суставе и морфологически сходными с тельцами Руффини и рецепторами Гольджи. 714 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Проприоцептивное восприятие положения тела и движений происходит в результате объединения в соматосенсорной коре информации от всех разновидностей проприоцепторов. Порог воспринимаемых изменений углового положения проксимальных суставов ниже, чем дистальных: 0,2— 0,4" в плечевом суставе и около Г в суставах пальца руки. Порог возрастает при увеличении угловой скорости движений, в связи с чем уменьшается точность выполняемых движений. Способность человека дифференцировать силу произвольных сокращений мышц подчиняется закону Вебера— Фехнера, эта способность уменьшается при самых слабых и очень сильных мышечных усилиях, необходимых для выполнения работы. 17.2.3. Температурная чувствительность Ощущения тепла или холода, вызванные прикосновением к телу человека нагретых или холодных предметов, теплого или охлажденного воздуха, возникают вследствие раздражения одной из двух разновидностей температурных рецепторов, представленных в коже свободными нервными окончаниями. Среди них различают тепловые и холодовые рецепторы, которым соответствуют две субмодальности температурного восприятия внешних воздействий. Температурные рецепторы распределены в коже неравномерно и образуют специфические тепловые и холодовые точки площадью около 1 мм2 и менее, с наибольшей плотностью на лице и ладонях, причем Холодовых точек примерно в десять раз больше, чем тепловых. Импульсная активность терморецепторов возрастает в зависимости от направления и скорости изменений температуры поверхности кожи, составляющей в условиях температурного комфорта около 33 °С. При ее снижении растет активность Холодовых рецепторов с максимумом в диапазоне от 26 до 17 °С, что субъективно расценивается как действие на кожу холода. При повышении температуры кожи от 34 до 45 "С увеличивается импульсная активность тепловых рецепторов, что субъективно ощущается как возрастающее по интенсивности действие на кожу тепла. При уменьшении температуры кожи ниже 17 °С и увеличении выше 45 °С к температурным ощущениям прибавляются болевые. Тоническая активность температурных рецепторов при неизменной температуре сохраняется в течение некоторого времени, а затем постепенно уменьшается. В таком случае, как и при медленных изменениях действующей на рецепторы температуры, происходит их адаптация, в связи с которой постепенно ослабевают ощущения тепла или холода. Но в ответ на резкие ступенчатые изменения действующей температуры (температурный контраст) возникает залповая фазическая активность терморецепторов, которая отражает произошедшие изменения температуры. Наличие адапта- 17. Сенсорные системы • 715
Кожные терморецепторы позволяют человеку распознавать нагретые или холодные предметы и окружающий воздух, прикасающиеся к коже. Однако субъективное восприятие, определяемое как «мне холодно» или «мне жарко», возникает в результате суммарной оценки информации, поступившей не только от кожных рецепторов, но также и от терморецепторов спинного мозга, внутренних органов и центральных рецепторов переднего гипоталамуса, где расположен центр терморегуляции. Нервные импульсы от кожных терморецепторов поступают в спинной мозг, в задних рогах которого расположены сенсорные нейроны второго порядка. Их аксоны переходят на противоположную сторону спинного мозга, входя в состав спиноретикулярного и спиноталамического путей. Спиноталамический путь ведет к задним вентролатеральным ядрам та-ламуса, на нейронах которых конвергируют проводящие пути температурной и тактильной чувствительности. Благодаря такой конвергенции у человека проявляется способность устанавливать, на какую часть поверхности тела подействовал температурный стимул. Спиноретикулярный тракт служит для проведения возбуждения от терморецепторов к ретикулярной формации и принадлежит к неспецифической части сенсорной системы. Ретикулярная формация вместе с неспецифическими ядрами таламуса активирует одновременно многие регионы коры, определяя общую активность мозга и уровень бодрствования. Поэтому повышение притока афферентной информации к ней от терморецепторов, например во время приема холодного душа, оказывает на человека тонизирующее влияние. 17.2.4. Болевая чувствительность Боль определяется как неприятное сенсорное и эмоциональное переживание, связанное с истинным или потенциальным повреждением ткани или описываемое в терминах такого повреждения. В отличие от других сенсорных модальностей боль всегда субъективно неприятна и служит не столько источником информации об окружающем мире, сколько сигналом повреждения или болезни. Болевая чувствительность побуждает к прекращению контактов с повреждающими факторами среды. Болевые рецепторы или ноцицепторы представляют собой свободные нервные окончания, расположенные в коже, слизистых оболочках, мышцах, суставах, надкостнице и во внутренних органах. Чувствительные окончания принадлежат либо безмякотным, либо тонким миелинизиро-ванным волокнам, что определяет скорость проведения сигналов в ЦНС и дает повод к различению ранней боли, короткой и острой, возникающей при проведении импульсов с большей скоростью по миелиновым волокнам, а также поздней, тупой и длительной боли, в случае проведения сигналов по безмякотным волокнам. Ноцицепторы относятся к полимодальным рецепторам, поскольку могут активироваться стимулами разной природы: механическими (удар, порез, укол, щипок), термическими (действие горячих или холодных предметов), химическими (изменение концентрации ионов водорода, действие гистамина, брадикинина и ряда других биологически активных веществ). Порог чувствительности ноцицепторов высок, поэтому лишь достаточно сильные стимулы вызывают возбуждение первич- 716 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Центральные отростки первичных сенсорных нейронов входят в спинной мозг в составе задних корешков и образуют синапсы с нейронами второго порядка, расположенными в задних рогах спинного мозга. Аксоны нейронов второго порядка переходят на противоположную сторону спинного мозга, где образуют спиноталамический и спиноретикулярный тракты. Спиноталамический тракт оканчивается на нейронах нижнего заднелате-рального ядра таламуса, где происходит конвергенция проводящих путей болевой и тактильной чувствительности. Нейроны таламуса образуют проекцию на соматосенсорную кору: этот путь обеспечивает осознаваемое восприятие боли, позволяет определять интенсивность стимула и его локализацию. Волокна спиноретикулярного тракта оканчиваются на нейронах ретикулярной формации, взаимодействующих с медиальными ядрами таламуса. При болевом раздражении нейроны медиальных ядер таламуса оказывают модулирующее влияние на обширные регионы коры и структуры лимбиче-ской системы, что приводит к повышению поведенческой активности человека и сопровождается эмоциональными и вегетативными реакциями. Если спиноталамический путь служит для определения сенсорных качеств боли, то спиноретикулярный путь предназначен играть роль сигнала общей тревоги, оказывать на человека общее возбуждающее действие. Субъективную оценку боли определяет соотношение нейронной активности обоих путей и зависимая от нее активация антиноцицептивных нисходящих путей, способных изменить характер проведения сигналов от ноци-цепторов. В сенсорную систему болевой чувствительности встроен эндогенный механизм ее уменьшения путем регуляции порога синаптических переключений в задних рогах спинного мозга («ворота боли»). На передачу возбуждения в этих синапсах влияют нисходящие волокна нейронов серого вещества вокруг водопровода, голубого пятна и некоторых ядер срединного шва. Медиаторы этих нейронов (энкефалин, серотонин, норадрена-лин) тормозят активность нейронов второго порядка в задних рогах спинного мозга, чем уменьшают проведение афферентных сигналов от ноци-цепторов. Аналгезирующим (обезболивающим) действием обладают опиатные пептиды (дайнорфин, эндорфины), синтезируемые нейронами гипоталамуса, которые имеют длинные отростки, проникающие в другие отделы головного мозга. Опиатные пептиды присоединяются к специфическим рецепторам нейронов лимбической системы и медиальной области таламуса, их образование увеличивается при некоторых эмоциональных состояниях, стрессе, продолжительных физических нагрузках, у беременных женщин незадолго перед родами, а также в результате психотерапевтического воздействия или акупунктуры. В результате повышенного образования опиат-ных пептидов активируются антиноцицептивные механизмы и повышается порог болевой чувствительности. Баланс между ощущением боли и ее субъективной оценкой устанавливается с помощью лобных областей мозга, участвующих в процессе восприятия болевых стимулов. При поражении лобных долей (например, в результате травмы или опухоли) порог болевой чувствительности не изменяется и поэтому сенсорный компонент болевого восприятия сохраняется неизменным, однако субъективная эмоциональная оценка боли становится иной: она начинает восприниматься лишь как сенсорное ощущение, а не как страдание. 17. Сенсорные системы • 717 17.2.5. Висцеральная чувствительность Большая часть афферентной информации от висцерорецепторов (у18сегаН8 — внутренностный) не осознается и служит для рефлекторной регуляции вегетативных процессов в организме человека. Висцерорецепторы расположены во внутренних органах и тканях организма и представляют собой как свободные, так и инкапсулированные нервные окончания. В зависимости от чувствительности к адекватным стимулам они подразделяются на меха-норецепторы, хеморецепторы, терморецепторы, осморецепторы и ноци-цепторы. Информация от висцерорецепторов поступает в ЦНС двумя путями: от сенсорных нейронов чувствительных ганглиев и по чувствительным афферентным волокнам, имеющимся в составе эфферентных симпатических и парасимпатических нервов. Висцеральные механорецепторы раздражаются вследствие изменения давления в полых органах и кровеносных сосудах. Осознаваемые ощущения, вызванные раздражением такого типа рецепторов, чаще всего обусловлены изменениями в пищеварительном тракте, поскольку из всех внутренних органов только он имеет общее эмбриологическое происхождение с поверхностью тела. Растяжение стенок желудка пищей вызывает ощущение сытости, растяжение стенок кишечника газами — ощущение дискомфорта и тяжести в области живота, а растяжение прямой кишки каловыми массами осознается как позыв к дефекации. Подобно этому им-пульсация от механорецепторов, расположенных в стенке мочевого пузыря, ощущается субъективно как позыв к его опорожнению. Висцеральные хеморецепторы обладают специфической чувствительностью к действию определенных химических молекул; например, хеморецепторы, расположенные в дуге аорты и синокаротидной области, раздражаются изменениями концентрации углекислого газа, кислорода и водородных ионов. Изменения активности таких рецепторов ощущаются не прямо, а опосредованно, проявляясь, например, головокружением или удушьем. Повышение осмотического давления вызывает раздражение периферических осморецепторов, расположенных в бассейне воротной вены, что, наряду с информацией от центральных осморецепторов промежуточного мозга, способствует появлению жажды. Афферентная информация от висцеральных терморецепторов служит для возникновения ощущений жара или озноба. Боль, возникающая при раздражении ноцицепторов внутренних органов, называется висцеральной, в отличие от соматической боли, вызванной раздражением болевых рецепторов в коже, мышцах, суставах и костях. Острую висцеральную боль может вызвать сильное растяжение полых органов вследствие закупорки выходных протоков желчного пузыря, мочеточников, в результате непроходимости кишечника, а также спастических сокращений гладких мышц. Плотность ноцицепторов особенно высока в брюшине, при воспалении которой или при ее сильном механическом раздражении возникает острая и интенсивная боль. При воспалительных процессах во внутренних органах выделяются биологически активные вещества, которые служат химическими раздражителями ноцицепторов и причиной болевых ощущений. 17.3. Зрительная сенсорная система Зрительное восприятие оставляет в памяти человека наибольшую часть его чувственных впечатлений об окружающем мире. Оно происходит в результате поглощения фоторецепторами сетчатки отраженной от окружающих 718 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Вкусовое восприятие Вкусовые ощущения отчетливо проявляются уже у новорожденных детей, а грудной ребенок 4—5 мес хорошо различает вкусовые изменения пищи. Вкусовое восприятие человека формируется в результате соединения муль-тимодальных сенсорных ощущений и механизмов памяти, хранящей индивидуальный опыт и связанной с усвоенными традициями питания. Изменения вкусового восприятия часто связаны с нарушениями обоняния, а не собственно вкусовой чувствительности. Ротовая полость сообщается с носовой, и пахучие вещества пищи достигают обонятельных рецепторов, поэтому представление о вкусе пищи почти всегда основано не только на вкусовом, но и на обонятельном восприятии. Гастрономические предпочтения людей связаны с комплексным действием тех или иных блюд, включающем не только вкусовые и обонятельные, но также тактильные температурные и зрительные ощущения. Переработка информации в переключательных ядрах и проводящих путях сенсорной системы Сенсорные нейроны низшего иерархического уровня способны передавать электрические сигналы одновременно нескольким нейронам следующего уровня благодаря дивергенции к ним своих аксонов. Это повышает надежность передачи информации от одного иерархического уровня к другому и позволяет сохранять чувствительность сенсорной системы при утрате отдельных нейронов. Возбуждение нейронов более высокого иерархического уровня определяется не только действием на них нейронов предыдущего уровня, но и механизмом латерального торможения, существующего в переключательных ядрах (рис. 17.2). Латеральное торможение происходит вследствие активации тормозных интернейронов переключательного ядра коллатералями возбуждающих нейронов. Чем сильнее возбужден релейный сенсорный нейрон, тем больше он активирует тормозные нейроны, которые подавляют активность соседних релейных нейронов. Релейные нейроны, передающие сигналы от центра рецептивного поля, сильнее других повышают активность тормозных нейронов, а их влияние на соседние релейные нейроны делает еще большим контраст между возбужденными и тормозимыми нейронами, выделяя линию передачи избранных сигналов. Высшие уровни сенсорной системы регулируют переработку информации в низших по отношению к ним переключательных ядрах посредством нисходящего торможения или усиления передаваемых сигналов. Нисходящее торможение (рис.17.3) происходит вследствие активации тормозных нейронов переключательного ядра, деятельность которых определяют нисходящие пути, начинающиеся на более высоком иерархическом уровне сенсорной системы. В результате нисходящего торможения повышается порог афферентной синаптической передачи в переключательном ядре низшего уровня. Нисходящее торможение представляет собой регуляцию сенсорно- 23* 708 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Рецепторы первичных сенсорных нейронов
Рис. 17.2. Схема латерального торможения в сенсорных системах. Тормозные интернейроны (их тела выделены черным цветом) возбуждаются возвратными коллатералями переключательных сенсорных нейронов: чем сильнее возбужден переключательный нейрон, тем большее торможение соседних переключательных нейронов он вызовет. Степень возбуждения переключательных нейронов определяется интенсивностью раздражения центра или периферии его рецептивного поля. В результате возбуждение будет передаваться от одной части рецептивного поля на фоне торможения или прерывания сигналов от другой его части, что создает эффект контрастирования, способствующий различению действующих стимулов. го восприятия по механизму отрицательной обратной связи. Тормозные интернейроны переключательных ядер уменьшают частоту передаваемых на следующий уровень сигналов, в связи с чем сенсорное ощущение ослабляется. Нисходящее усиление происходит с помощью возбуждающих интернейронов переключательного ядра, которые активируются аксонами нейронов более высокого иерархического уровня. Вследствие этого в управляемом по механизму положительной обратной связи переключательном ядре понижается порог синаптической передачи, что делает возможным прохождение относительно слабых сигналов на следующий иерархический уровень. Одновременное использование механизмов отрицательной и положительной обратной связи позволяет освобождаться от избыточной информации, подавлять «шум», т. е. нейронную активность, не содержащую физиологически значимых сообщений, и одновременно выделять и усиливать сигналы, на которых будет сосредоточено внимание. Большинство раздражителей, воспринимаемых сенсорными системами, оказывает на них комплексное действие, поскольку одновременно возбуждает разные типы рецепторов. Например, прикасающийся к ограниченному участку кожи предмет может быть гладким или шершавым, теплым или холодным, сухим или влажным, а его давление на кожу может усиливаться или ослабевать, он может передвигаться в том или ином направлении. 17. Сенсорные системы • 709
сенсорной системы К другим областям ЦНС Афферентные сигналы Рис. 17.3. Схема нисходящего торможения в сенсорных системах. Передачу афферентных сигналов от одного иерархического уровня сенсорной системы к другому в переключательных ядрах контролируют высшие иерархические уровни. На схеме показан принцип нисходящего торможения коры, которое осуществляется с помощью тормозных интернейронов (выделены черным цветом) переключательного ядра. Возбуждение тормозных интернейронов под влиянием коры препятствует передаче афферентных сигналов в переключательном ядре. Зрительные ощущения представляют собой сочетание определенной формы воспринимаемого предмета, его движения и комбинации различных цветов, заполняющих ту или иную деталь формы. Сенсорные системы организованы так, что информация о каждом компоненте комплексного раздражителя передается одновременно по нескольким параллельным путям, каждый из которых относится к определенной субмодальности, как, например, форма, движение или цвет наблюдаемого зрительного объекта. В каждом из таких путей независимо друг от друга перерабатываются сигналы, несущие информацию о разных качествах подействовавшего на рецепторы комплексного раздражителя. При этом разные каналы, используемые для передачи сенсорной информации, не дублируют друг друга, в чем проявляется принцип многоканальности, присущий всем сенсорным системам. Высокая упорядоченность нейронных переключений на каждом иерархическом уровне обеспечивает поступление информации от каждого рецептивного поля к соответствующим именно ему кортикальным колонкам проекционной коры. Необходимое для целостного восприятия раздражителя объединение информации, относящейся к разным субмодальностям, происходит в сенсорных регионах коры. 710 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Каждая сенсорная система воспринимает действие адекватного стимула в ограниченном диапазоне значений его силы. Наименьший по интенсивности стимул, способный вызвать ощущение, называется абсолютным порогом ощущения. Его величина устанавливается опытным путем, она не одинакова у разных людей и может изменяться у одного и того же человека в зависимости от функционального состояния, различаясь, например, при оптимальной работоспособности и утомлении. Величина абсолютного порога ощущения выше абсолютного порога первичных сенсорных нейронов, так как возбуждение отдельных чувствительных нейронов или их небольшой группы не обязательно приводит к возбуждению высших уровней сенсорной системы вследствие тормозных процессов на предыдущих иерархических уровнях. Поэтому не каждый стимул, возбуждающий рецепторы первичного сенсорного нейрона, субъективно ощущается и осознается. Величина, на которую один стимул надпорогового диапазона должен отличаться от другого, чтобы их разницу можно было субъективно различить, получила название дифференциального порога или порога различения. Ощущаемый прирост интенсивности раздражения (дифференциальный порог) должен превышать ранее действовавший стимул на определенную и постоянную величину. Например, человек легко различает по весу гирьки весом 14 и 15 граммов (разница в 1 грамм), но гирьки весом 29 и 30 граммов кажутся ему одинаковыми, и различия между ними выявляются, если разница окажется не меньше 2 граммов. Если же гирька составит 60 граммов, то субъективное ощущение большего груза возможно только тогда, когда его прирост будет не меньше 4 граммов. Таким образом, ощущаемый прирост силы раздражителя возможен при ее увеличении на постоянную величину, что сформулировано в законе Вебера как 68/8 = к, где 3 — раздражение, 63 — его ощущаемый прирост, к — постоянная величина. Закон Вебера справедлив для разных модальностей, но лишь в границах раздражения средней силы, а при слабых и сильных стимулах способность к различению силы стимулов у человека снижается. Предложена также ступенчатая шкала интенсивности ощущений, в которой за ноль принимается величина абсолютного порога, а зависимость между интенсивностью стимула и ощущением выражается как: Е = к х 1о§(8/80), где Е — интенсивность ощущения, к — постоянная величина, 3 — интенсивность стимула, 80 — абсолютный порог. Эта зависимость, получившая название закона Вебера—Фехнера, показывает, что линейное увеличение интенсивности ощущения отражает логарифмический рост интенсивности стимула. Закон Вебера—Фехнера характеризует субъективную способность человека различать действующие стимулы (шкала различения), но не оценивает саму интенсивность ощущений, поскольку этот закон основан на допущении равенства любых ее приростов, как к слабым, так и к сильным стимулам. Субъективная оценка интенсивности раздражителя (качественное различие) возрастает при увеличении разницы между пороговой и действующей силой стимула (количественное различие). Однако зависимость между силой стимула и ощущением не одинакова при разной интенсивности стимулов, а потому имеет не линейный, а степенной характер. Для оценки интенсивности ощущений всего диапазона используется шкала Стивенса, устанавливающая зависимость ощущений от силы стимула в виде степенной функции: Е = к(8 — 80)п, где Е — интенсивность ощущения, 3 — действующая сила стимула, 30 — абсолютный порог, к — константа шкалы, п — показатель степени, который зависит от сенсорной модальности (на- 17. Сенсорные системы • 711
Пространственные характеристики действующих стимулов, необходимые для их различения, зависят от специфических особенностей каждой сенсорной системы и величины рецептивных полей. Прикосновение к коже дистальной фаланги пальца руки двух ножек циркуля с расстоянием между ними 2 мм ощущается раздельно, но чтобы ощутить раздельное прикосновение к коже спины, ножки циркуля необходимо раздвинуть до 60 мм. Пространственное восприятие этих тактильных стимулов зависит от размеров соответствующих рецептивных полей: раздельное ощущение возможно только при условии раздражения каждой ножкой циркуля независимого рецептивного поля. Лишь тогда информация о каждом стимуле будет перерабатываться раздельно на каждом уровне организации сенсорной системы, включая проекционную область коры. Аналогичная ситуация имеет место при восприятии двух точек зрительного поля: они не сливаются в одну, если отражаемые ими световые лучи попадут на разные рецептивные поля сетчатки. Имеет значение и степень контраста между действующим стимулом и его фоном: хорошо контрастируемые объекты (например, черное на белом) различаются легче, чем мало контрастируемые (черное на сером). Временная характеристика восприятия действующих стимулов у человека имеет абсолютный порог различения коротких временных отрезков, который соответствует примерно '/18 секунды. Например, 18 зрительных изображений, предъявленных в течение 1 секунды, сливаются в непрерывное движение, 18 прикосновений к коже за 1 секунду воспринимаются как одно, а 18 звуковых колебаний в секунду воспринимаются как один очень низкий звук. Разрешающая способность сенсорных систем для восприятия действующих через малые промежутки времени стимулов ограничена рефрактерным периодом, во время которого сист
|
||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-21; просмотров: 393; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.135.249.76 (0.013 с.) |


 чания волосяных фолли
чания волосяных фолли
 Афферентная информация от проприоцепторов в составе задних корешков поступает в спинной мозг и без переключения в нем передается по восходящим путям к ядрам задних столбов продолговатого мозга. Расположенные там сенсорные нейроны второго порядка передают информацию по лемнисковому пути на противоположную сторону мозга к проекционным ядрам таламуса. Нейроны переключательных ядер таламуса образуют соматотопическую проекцию на соматосенсорную кору, с помощью которой формируются проприоцептивные ощущения. Сенсорные сигналы от проприоцепторов мышц лица и головы следуют в ЦНС по чувствительным волокнам черепных нервов, а в соматосенсорную кору информация поступает тем же способом, что и сигналы от мускулатуры тела, т. е. по лемнисковому пути.
Афферентная информация от проприоцепторов в составе задних корешков поступает в спинной мозг и без переключения в нем передается по восходящим путям к ядрам задних столбов продолговатого мозга. Расположенные там сенсорные нейроны второго порядка передают информацию по лемнисковому пути на противоположную сторону мозга к проекционным ядрам таламуса. Нейроны переключательных ядер таламуса образуют соматотопическую проекцию на соматосенсорную кору, с помощью которой формируются проприоцептивные ощущения. Сенсорные сигналы от проприоцепторов мышц лица и головы следуют в ЦНС по чувствительным волокнам черепных нервов, а в соматосенсорную кору информация поступает тем же способом, что и сигналы от мускулатуры тела, т. е. по лемнисковому пути. ции терморецепторов наряду с их способностью реагировать залповой активностью на контрастные изменения действующей температуры проявляются в уменьшении порога тепловой чувствительности при пониженной температуре кожи и в уменьшении порога холодовой чувствительности при повышении ее температуры.
ции терморецепторов наряду с их способностью реагировать залповой активностью на контрастные изменения действующей температуры проявляются в уменьшении порога тепловой чувствительности при пониженной температуре кожи и в уменьшении порога холодовой чувствительности при повышении ее температуры. ных сенсорных нейронов: например, порог болевой чувствительности для механических стимулов примерно в тысячу раз превышает порог тактильной чувствительности.
ных сенсорных нейронов: например, порог болевой чувствительности для механических стимулов примерно в тысячу раз превышает порог тактильной чувствительности. предметов энергии световых лучей или электромагнитных волн в диапазоне от 400 до 700 нм. Энергия поглощенных квантов света (адекватный раздражитель) преобразуется сетчаткой в нервные импульсы, поступающие по зрительным нервам к латеральным коленчатым телам, а от них — в проекционную зрительную кору. В дальнейшей переработке зрительной информации у человека участвуют свыше тридцати отделов мозга, представляющих вторичные сенсорные и ассоциативные области коры.
предметов энергии световых лучей или электромагнитных волн в диапазоне от 400 до 700 нм. Энергия поглощенных квантов света (адекватный раздражитель) преобразуется сетчаткой в нервные импульсы, поступающие по зрительным нервам к латеральным коленчатым телам, а от них — в проекционную зрительную кору. В дальнейшей переработке зрительной информации у человека участвуют свыше тридцати отделов мозга, представляющих вторичные сенсорные и ассоциативные области коры. Действие раздражителя
Действие раздражителя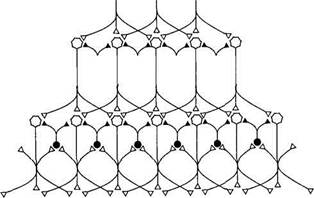
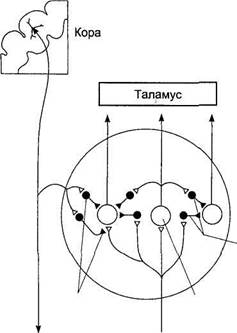 Переключательное ядро
Переключательное ядро


