Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Понимание и центральное представление пространстваСодержание книги
Поиск на нашем сайте
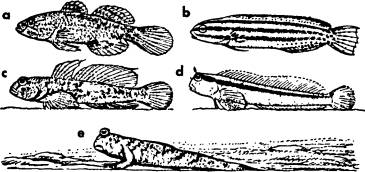
"Понимающее" поведение и «разумность» тесно связаны друг с другом: разумным считается существо с высокоразвитой способностью понимающего поведения. О том, как эта способность возникает, старая психология, очевидно, мало задумывалась — может быть, потому, что эту в высшей степени духовную функцию заранее считали физиологически необъяснимой. Поэтому определение способности к понимающему поведению ограничивается обычно отрицательными положениями. Как уже было указано (с. 266), форма поведения считается понимающей, если она позволяет организму справляться с некоторой специальной ситуацией окружающей среды способом, содействующим сохранению вида, хотя об этой особенной ситуации организм не располагает ни информацией, приобретенной в ходе эволюции, ни полученной в индивидуальной жизни. Как мы уже говорили в 1.3 и подробнее во всей главе 4, с этим чисто отрицательным определением "per exclusionem"[49] можно сопоставить другое, положительного типа, основанное на знании механизмов, лежащих в основе понимающего поведения: мы называем понимающей форму поведения, особая приспособленность которой опирается на процессы кратковременного получения информации, описанные в главе 4. В ориентации высших позвоночных особо важную роль играет оптическое восприятие пространственной глубины и направления. Поэтому даже наивный наблюдатель не совсем неправ, когда он судит о разумности отдельных видов животных по более высокому или более низкому развитию у них этих способностей. Между тем есть большое число позвоночных, получающих информацию для пространственной ориентировки главным образом из вызываемых собственным движением животного параллактических[50] смещений изображений отдельных предметов на сетчатке. Этот вид пространственного восприятия наблюдается не только у рыб, но и у бесчисленных птиц и млекопитающих; достаточно вспомнить зуйка или малиновку, вынужденных совершать для локализации видимого предмета характерные кивающие или покачивающиеся движения. Копытные также лишь в исключительных случаях фиксируют предметы обоими глазами, и даже собака, бегущая без поводка рядом с хозяином, поддерживая с ним при этом тесный контакт, очень редко смотрит на него обоими глазами, разве что хозяин позовет ее, чихнет, споткнется и т. п. и этим привлечет ее внимание. Но даже в таком случае собака чаще всего смотрит на хозяина, держа голову наискось, что указывает на преимущественно акустическую ориентировку. Преобладание у столь многих высших животных параллактической ориентации приводит, как можно заметить, многих людей к ошибочному мнению, что все эти животные не в состоянии фиксировать предметы обоими глазами. Поэтому нужно особо подчеркнуть: всем двуглазым позвоночным в принципе присущ механизм ориентации, состоящий в том, что изображение предмета на сетчатке приводится посредством двустороннего направления глаз в положение наиболее резкой видимости, а затем по обратным сообщениям глазных мышц определяется расстояние до этого предмета. У немногих видов, не обладающих таким механизмом, он, вероятно, вторичным образом атрофировался, как, например, у ряда сомов (Siluridae) вьюновых (Cobitidae), и в немногих других случаях. Весьма вероятно, что зрение двумя глазами вообще возникло под селекционным давлением функции бинокулярной фиксации добычи. У многих рыб, земноводных и пресмыкающихся, у которых, за исключением случаев бинокулярной фиксации, глаза движутся независимо друг от друга, пеленгация[51] добычи выполняется примерно следующим способом. Сначала животное фиксирует добычу одним глазом, внимательно следящим за всеми ее движениями. Когда интенсивность реакции возрастает, второй глаз также поворачивается к добыче. После этого животное устанавливает свою голову или все тело в плоскости симметрии между направлениями зрения обоих глаз, неизменно направленных на добычу, а затем приближается к добыче до тех пор, пока достигается надлежащее расстояние, чтобы ее схватить. При этом информацию о величине фиксируемого предмета и расстоянии до него доставляют, по всей вероятности, те же механизмы, которые Эрих фон Гольст обнаружил у людей. Как конвергенция[52] глазных осей, так и процесс установки хрусталиков на резкость доставляют путем реафференции необходимые данные. Многие животные с особенно дифференцированным механизмом ловли добычи не способны схватить ее и в том случае, если она находится слишком близко от их рта. Например, морской конек вынужден при этом с трудом отодвигаться, часто изгибая гротескным образом свое тело, чтобы маленький рачок, неудобно подплывающий к нему, оказался в плоскости симметрии его головы и в то же время на подходящем "расстоянии выстрела". У значительного большинства позвоночных оценка параллактического смещения видимых предметов служит для грубой ориентировки, а бинокулярное зрение — для локализации добычи. Бинокулярная фиксация неживых окружающих предметов, имеющих значение лишь как препятствия или субстрат перемещения,[53] происходит лишь в тех особых случаях, когда такой предмет должен быть точно локализован, например, когда в ходе движения нога животного должна быть точно поставлена на данный предмет или когда хватательный орган должен его с уверенностью охватить. С такими требованиями сталкиваются уже некоторые рыбы. а именно живущие на дне или ползающие формы, потерявшие способность свободно парить в воде; так как они тяжелее воды, они остаются на дне, если только не поднимаются над ним усиленными плавательными движениями. Когда таким донным формам приходится целенаправленно перемещаться в богато структурированной среде, например в скалистых нагромождениях литорали[54] или в зарослях мангровых корней, они должны быть способны точно локализовать детали поверхности, по которой ползают. Несколько групп окуневых рыб (Percomorphae) выработали формы, специализировавшиеся на этом. Особенно хорошо оснащены в этом смысле представители собачковых (Blenniidae) и пескарей (Gobiidae). Почти все наблюдатели этих интересных рыбок подчеркивают их «разумность». Уильям Биб говорит о них: "Of all fishes, they are least bound-up in fishiness",[55] а мой учитель профессор Генрих Йозеф говорил о них в шутку, но очень метко: "Blennius вообще не относится к классу рыб, а принадлежит к таксам". Этим хорошо описывается комическое впечатление, всегда возникающее у опытного наблюдателя, когда животное делает нечто неожиданное, особенно то, что мы привыкли видеть лишь у более высокоразвитых животных. Когда мы видим, как бленниус, взобравшийся "собственными ногами" на высокий камень, затем высоко поднимается на брюшных плавниках, загнутых далеко вперед, сильно отклоняет вверх голову — почти у всех других рыб жестко связанную с туловищем — и фиксирует обоими глазами верхний край обломка скалы, прежде чем вскочить на него точно нацеленным прыжком, — все это каждый раз производит неотразимо комическое впечатление. Еще забавнее Periophthalmus, принадлежащий к Gobiidae, который в фиксации обоими глазами неживых предметов и в ползании превосходит всех Blenniidae. Он способен, выйдя из воды, взбираться на мангровые корни и целенаправленно перепрыгивать с одного из них на другой. Среди Blenniidae и Gobiidae есть и первоначальные формы, не дифференцированные в указанном направлении, с вполне дееспособным плавательным пузырем, так что они, подобно другим рыбам, могут свободно парить в воде без мышечных усилий. При сравнении их с ползающими формами той же группы бросается в глаза существенное различие в форме головы. У способных к парению форм лоб плосок, как и у других рыб, и глаза расположены по бокам головы. Чем дальше заходит приспособление к жизни на дне и к ползанию и чем важнее становится тем самым функция точной оптической ориентации в пространстве, тем круче становится лоб и тем ближе подходят глаза к грани между спинной линией и передней стороной головы, так что перед ними открывается свободное пространство для бинокулярной фиксации. Это приспособление достигает крайней степени у уже упомянутого илистого прыгуна,[56] Periophthalmus. На рис. 2 показана последовательность приспособления у различных видов обоих порядков. Сами по себе оценка параллактического смещения видимых предметов и локализация их бинокулярной фиксацией — это два различных, но вполне равноценных механизма. И все же антропоморфная точка зрения, по которой фиксирующие животные умнее животных с неподвижным взглядом, не совсем ошибочна. Виды рыб, лучше всех известные человеку, как, например, золотые рыбки и многие другие, имеют вошедшие в поговорку неподвижные "рыбьи глаза". Все они принимают пространственно ориентирующую информацию, лишь когда движутся, поскольку лишь в таком случае изображения на сетчатке различных предметов пространства смещаются по отношению друг к другу. Когда такая рыба останавливается вплотную перед каким-нибудь препятствием, например камнем или плотным кустом водяного растения, то часто можно заметить, что, когда она снова начинает двигаться, она вначале продолжает в точности следовать направлению, прегражденному препятствием. Лишь когда собственное движение вызывает соответствующий сдвиг изображений на сетчатке и тем самым доставляет информацию, лежащую в основе ориентации, рыба начинает изменять направление плавания. Поведение животных, ориентирующихся с помощью фиксации, отличается от только что описанного в одном существенном отношении: в то время как при параллактической ориентации перемещение и пеленгация, как мы видели, совпадают во времени, у фиксирующих животных они разделены во времени. Локализация предшествует перемещению. Животные, ориентирующиеся с помощью «телотактической» фиксации, т. е. приводящие изображения предметов одно за другим в место острейшего зрения, как бы ощупывая пространство вокруг себя, всегда делают это, оставаясь на месте. Вслед за этой фазой ориентации начинается перемещение, уже вполне ориентированное и как будто спланированное заранее. Аналогии с этой последовательностью внутреннего приготовления и последующего выполнения движения встретятся нам еще раз на более высоких уровнях функций понимания.
Рис. 2. Конвергентное приспособление положения глаз к бинокулярной фиксации субстрата у Gobiidae и Blenniidae. Свободно плавающие формы, Dormitator, порядок Gobiidae, (а) и Petroscirtes, порядок Blenniidae, (b) имеют такую же форму головы, как большинство свободно плавающих рыб. Обитающие на дне формы, такие, как Gobius jozo (с) и Blennius rouxi (d), имеют крутой профиль головы, делающий возможной сильную конвергенцию глаз вперед. У живущего на суше Periophthalmus, порядок Gobiidae (e) это приспособление заходит дальше всего.
Конечно, параллактическая и фиксирующая пространственная ориентация встречаются вместе друг с другом, в одном и том же организме; роли их различаются лишь количественно. Для параллактической ориентации важно периферическое зрение, т. e. оценка данных, доставляемых более далекими от центра сетчатки местами; поэтому у животных, способных к хорошей параллактической ориентации, глаза часто бывают больше, чем у постоянно прибегающих к фиксации. Например, чиж непрерывно фиксирует во всех направлениях "все, что есть в пространстве", и его крохотные глазки находятся в постоянном движении. Между тем большой глаз малиновки или зуйка остается почти неподвижным относительно головы, но зато через правильные промежутки времени производится уже упомянутое покачивание. Примечательным образом два способа ориентации во многих случаях не могут заменять друг друга. Как уже было сказано, первичная функция бинокулярного восприятия глубины — прицеливание в добычу, тогда как параллактическое восприятие глубины служит для грубой ориентации в пространстве и избежания препятствий. Рыбы открытого моря, обладающие как хищники превосходным бинокулярным зрением, не в состоянии применять его для избегания препятствий. На Адриатическом побережье мне довелось однажды видеть, как сотни мальков одного вида морских щук (Belonidae)[57] плыли прямо на берег и там гибли. Они двигались не сомкнутой стаей, а поодиночке, на расстоянии около метра друг от друга и в точности параллельно, очевидно управляемые какой-то ориентировкой на свет, соленость воды или на что-то еще. За несколько метров от берега все они были еще вполне здоровы, в зоне прибоя они боролись за свою жизнь, а на берегу нагромождался вал из трупов. О рыбах открытого моря, живущих в неволе, также известно, что они не в состоянии принимать во внимание препятствия, расположенные под прямым углом к направлению плавания, а потому даже в очень больших бассейнах сплошь и рядом наталкиваются головой на стенку. В открытом море они обнаруживают уже издали большие и непрозрачные предметы с помощью параллактической ориентации, обходя их по плавным дугам. С ситуацией, когда подобные препятствия окружают их со всех сторон, они не в состоянии справиться. Тесная корреляция между способностью к пространственной ориентации и пространственной структурой жизненного пространства уже знакома нам на примере приспособления Gobiidae и Blenniidae к сухопутной жизни и ползанию. Та же корреляция существует и вообще во всем животном мире. Меньше всего понимание требуется от организмов, живущих в однородной по всем направлениям среде — в открытом море. Из всех известных нам в настоящее время свободно движущихся многоклеточных животных самые «глупые» — это, по-видимому, некоторые медузы, которые, как, например, легочная медуза (Rhisostoma pulmo),[58] вообще лишены пространственно направленных реакций. Они и не нуждаются в таковых, поскольку их легкий колокол и тяжелая желудочная ножка удерживают их в устойчивом равновесии, а пищу они добывают фильтрацией из морской воды, так что им не надо схватывать целенаправленным движением отдельные частицы еды. Единственные встречающиеся у них направленные движения — это движения амебоидных клеток стенок желудка. Единственный ответ всего организма на стимулы состоит в том, что каждый удар зонтика возбуждает определенные рецепторы, так называемые краевые тельца, чем запускается следующее сжатие этого органа. Как остроумно сказал о легочной медузе Якоб фон Юкскюль, "она не воспринимает ничего, кроме ударов собственного колокола". Как мы уже знаем, многие рыбы открытого моря не способны целесообразно реагировать на пространственные условия. У сухопутных животных также существует тесная связь между структурированностью жизненного пространства и способностью справляться с нею при помощи пространственно ориентированного поведения. Открытую степь можно в некотором смысле считать двумерным аналогом открытого моря, однородного во всех трех пространственных направлениях. Если, например, сравнить способность к пространственному пониманию у близко родственных форм животных, одни из которых живут в степях, а другие — в скалах или на деревьях, скажем, у куропатки и хохлатого перепела или у антилопы орикс и серны, то обнаруживаются поразительные различия в способности овладения пространственными условиями. Пара куропаток, воспитанных мною "с яйца", оказалась не способной понять как непроницаемое препятствие выбеленную стену. Когда им хотелось переменить место, они бежали в направлении стены комнаты, расположенной против окна, потому что она была светлее, чем часть стены под окном. Встретившись с препятствием, они не просто натыкались на него клювом, а упорно продолжали толкаться в том же направлении, неустанно отбегая и снова набегая на стену. Повернувшись, они с особенной силой мчались вперед, все больше стирая себе роговую поверхность кончика клюва и перья передней части шеи, так что мне пришлось повесить перед стеной темный занавес, чтобы предотвратить более серьезные повреждения. Чтобы ограничить птиц определенной частью комнаты, я отделил их доской, вышины которой едва хватало, чтобы не позволить им выглядывать наружу. Куропатки часто налетали на эту доску то выше, то ниже, но так и не научились целенаправленно перелетать через нее даже после того. как они много раз при бесцельных взлетах случайно оказывались по другую сторону доски. Примечательно, что в полете они гораздо лучше ориентировались в пространстве, чем на земле. Когда они внезапно взлетали с большим ускорением, на которое эти птицы способны, казалось, что сейчас раздастся сильный удар о стену или оконное стекло и на пол упадет мертвая птица. Но этого никогда не происходило; куропатки всегда благоразумно сворачивали, оказавшись вблизи стены, и целенаправленно приземлялись на полу. Ясно, что куропатка может "позволить себе" пренебрегать препятствиями на степной почве, но в полете должна быть в состоянии избегать таких вертикальных препятствий, как дерево, опушка леса или крутой склон. Передвигаясь на ногах, куропатка так же не может использовать свои способности к ориентации, «предусмотренные» для полета, как морская щука не может использовать свое «предусмотренное» для ловли добычи бинокулярное зрение для того, чтобы не натолкнуться на крутой скалистый берег. Как уже было сказано, способности к овладению сложными пространственными условиями часто сильно различаются у очень близко родственных форм. У морских щук, которые не в состоянии даже «понять» как препятствие крутой берег, есть близкие пресноводные родственники, каким-то образом справляющиеся даже со стеклянными стенами маленького аквариума, хотя, как это видно на многих фотографиях, большей частью ценой повреждения своих острых челюстей. Хохлатый перепел, близкий родственник куропатки, отлично ориентируется в пространстве, и т. п. Так как вряд ли можно допустить, что у этих животных органы чувств устроены иначе, чем у их "более глупых" родственников, можно предположить, что способность к лучшему или худшему представлению пространства зависит от организации центральной нервной системы. Какое же жизненное пространство предъявляет к своим обитателям наивысшие требования в отношении понимания пространства? Без сомнения, это ветви деревьев! Но среди древесных животных самая точная, доходящая до мельчайших деталей информация о пространственных условиях требуется не тем, которые удерживаются на ветвях с помощью присосок или когтей, как это делаю ι древесные лягушки, белка и многие другие обитатели деревьев; она нужна тем, кто лазает по деревьям с помощью хватательных органов, сжимающих как клещи сучья и ветки. Животные, лазающие с помощью присосок и когтей, решаются броситься приблизительно в направлении намеченной цели; вероятность уцепиться за что-нибудь одним или двумя пальцами довольно велика, да и падение у этих большей частью маленьких животных не обязательно приводит к трагическому концу. Напротив, хватающая рука дает опору лишь в том случае, если она сжимается в надлежащем месте, в надлежащем положении и в надлежащий момент; она не прикрепляется ни когда она раскрыта, ни когда сжата в кулак. Хватание сжимающей рукой коррелировано у млекопитающих с определенным положением глаз на голове аналогично тому, как у рыб коррелирована с положением глаз редукция плавательного пузыря и способности к парению, и по аналогичным причинам. В обоих случаях требуется особенно точное представление пространства. Среди сумчатых, как и среди плацентарных, имеются обитатели деревьев, лазающие с помощью когтей и с помощью хватающих рук. В обоих подклассах млекопитающих первые имеют большей частью расположенные по бокам и несколько выпученные глаза, как мы это видим у белки, между тем как вторые имеют большей частью глаза, направленные вперед, "как у совы". У сумчатых рассматриваемая корреляция менее выражена, вероятно, потому, что многие их формы лазают очень медленно и не нуждаются при своем хамелеонообразном перемещении в особенно хорошей пространственной ориентации. Но можно с уверенностью утверждать, что все животные, захватывающие свою цель после далекого прыжка клещеобразной рукой, предварительно фиксируют эту цель бинокулярным зрением. Висение и лазание антропоидов с их крюковидной рукой предъявляет отнюдь не меньшие требования к точности пространственного понимания,[59] чем у других приматов с хватающей рукой. Но только крюковидная рука обладает особым свойством, к которому мы еще вернемся в разделе о любознательном поведении и самоисследовании: во время захвата она находится в непосредственной близости от точки фиксации. При описании поведения животных, ориентирующихся посредством фиксации различных окрестных предметов, я уже сказал, что временная последовательность движений глаз, «прощупывающих» пространство, и заключительного уверенного и успешного движения производят впечатление чрезвычайной «разумности»; причина этого в том, что такое поведение есть простой аналог планирующего мышления, какое мы находим у человекообразных обезьян. Когда антропоиду предлагают задачу, требующую понимания, он ведет себя совсем иначе, чем енот или макака-резус в подобном случае. Те беспокойно бегают в поиске взад и вперед, моторно испытывая разные возможности. Но человекообразная обезьяна спокойно усаживается, внимательно обводя взглядом экспериментальную установку. Ее внутренняя напряженность выражается в так называемых проскакивающих движениях, например очень часто она почесывает себе голову, как размышляющий человек. Она также «испытывает» разные возможности, на что указывает блуждание ее взгляда, без устали перепрыгивающего с одной точки экспериментальной установки на другую. Это очень хорошо видно в фильме об опытах с орангутаном, снятом в Сухуми. в Советском Союзе. Перед обезьяной поставили задачу передвинуть ящик, стоящий в одном из углов помещения, в противоположный угол, где к потолку на нитке был подвешен банан. Вначале взгляды обезьяны беспомощно блуждают по диагонали между ящиком, стоящим в левом нижнем углу, и бананом, висящим справа вверху. Затем обезьяна начинает злиться, так как не может найти решение; она пытается уклониться от неприятного положения, отвернувшись — cut-off behaviour[60] в смысле Чанса. Но задача не дает ей покоя, она снова поворачивается к экспериментальной установке. Вдруг ее взгляд начинает двигаться другими путями. Он обращается к ящику, затем к месту на полу под бананом, оттуда вверх к манящей цели, снова вертикально вниз и опять к ящику. После этого молниеносно следует освобождающая, решающая задачу догадка, которую можно безошибочно прочесть на выразительном лице оранга, и он тотчас же направляется, кувыркаясь от радости, к ящику, подставляет его под банан и достает его. Для необходимого понимающего поведения ему было достаточно нескольких секунд. Кто наблюдал такое решение задачи обезьяной, не может всерьез усомниться, что в момент решения животное испытывает переживание, аналогичное нашему "переживанию «ага» в смысле Карла Бюлера. Что же происходит с обезьяной объективно и субъективно, когда она неподвижно сидит, но совершает при этом трудную внутреннюю работу, собирая взглядом информацию о заданной ситуации? Что она переживает, мы не знаем, но мы можем с большой уверенностью допустить, что весь этот процесс аналогичен тому, который мы у самих себя называем мышлением. Лично я уверен в том, что обезьяна делает то же, что и я сам, а именно в воображаемом, т. е. модельно представленном в ее центральной нервной системе, пространстве передвигает столь же воображаемый ящик и "представляет себе", как она затем сможет на него взобраться и достать банан. Я не вижу, как может быть мышление чем-то принципиально иным, чем такое пробное, совершающееся лишь в мозгу действие в воображаемом пространстве. Я утверждаю, по меньшей мере, что процессы этого рода содержатся также в наивысших операциях нашего мышления и составляют их основу. Во всяком случае, я не в силах представить себе какую-нибудь форму мышления, независимую от этой основы. Уже более 20 лет назад В. Порциг написал в своей книге "Чудо языка" следующие слова: "Язык переводит все не наглядные отношения в пространственные. И это делает не какой-то один язык или группа языков. а все без исключения. Эта особенность принадлежит к неизменным чертам ("инвариантам") человеческого языка. Временные соотношения выражаются в языке в пространственных терминах: перед Рождеством и после[61] Рождества, на протяжении двух лет.[62] Когда речь идет о психических процессах, мы говорим не только о внешнем и внутреннем, но также о том, что происходит "выше и ниже порога" сознания, о подсознании, о переднем плане и заднем плане, о глубинах[63] и слоях психики. Пространство вообще служит моделью для всех не наглядных отношений: наряду[64] с работой он преподает; любовь была больше честолюбия; за[65] этой мерой стоял замысел — незачем приводить дальнейшие примеры, любое число которых можно найти в каждом куске письменной или устной речи. Значение этого явления связано с его всеобщей распространенностью и его ролью в истории языка. Его можно продемонстрировать не только на примере употребления предлогов, которые все первоначально означают пространственные отношения, но также на словах, обозначающих виды деятельности и свойства". Уже в работе, вышедшей в 1954 году, я выразил ту точку зрения, что приведенные выше выводы языковеда "имеют основополагающее значение не только для истории языка, но еще более для филогенетического развития мышления вообще и тем самым также дословесного и несловесного мышления". Эта точка зрения получила в последнее время в высшей степени важное подтверждение в исследованиях Ноама Хомского и его сотрудников (4). На основе обширного сравнительного изучения языков эти исследователи пришли к выводу, что определенные основные структуры языка и мышления свойственны всем людям всех культур и являются врожденными, причем в одинаковой форме. По их мнению, эти общечеловеческие и специфические для нашего вида функции возникли не под селекционным давлением взаимопонимания, а под селекционным давлением логического мышления. Независимо от Хомского, Герхард Гёпп пришел к следующему выводу, изложенному в его книге "Эволюция языка и разума": "Язык есть не только средство внешнего взаимопонимания, но и неотъемлемая составная часть самого разума".
|
||||
|
|
Последнее изменение этой страницы: 2024-06-27; просмотров: 5; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.189.141.236 (0.015 с.) |