Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Чувствительные восходящие пути.Содержание книги
Поиск на нашем сайте
Все заднекорешковые волокна и вообще все волокна, идущие от рецепторов, кроме обонятельных, связаны с новой корой больших полушарий, но ни одно из них не достигает ее без перерыва. Первоначально волокна прерываются где-либо в спинном мозге или в ядрах ствола головного мозга и мозжечка, а отсюда аксоны новых нейронов направляются к коре. Но они в большинстве случаев прерываются в зрительных буграх. Некоторые нейроны второго порядка оканчиваются в четверохолмии, например, нейроны слухового и зрительного нервного пути. Все спинномозговые чувствительные пути проходят сначала через петлю Рейля, а затем через зрительные бугры. Таким образом, восходящие корковые пути должны прерываться минимум дважды, а потому состоят, по крайней мере, из трех нейронов. Большая часть чувствительных восходящих путей перекрещивается. Неперекрещивающиеся пучки существуют в восходящих путях от кожномышечных и зрительных рецепторов. Совсем не перекрещиваются обонятельные пути, а слуховые пути перекрещиваются полностью. Перекрест обычно происходит в области второго нейрона. Так аксоны заднекорешкового первого афферентного нейрона перекрещиваются в пути от кожно-мышечных рецепторов, а аксоны третьего нейрона, кончаю- щиеся в коре, не перекрещиваются. Так как часть заднекорешковых волокон кончается в спинном мозге, а часть в продолговатом, то и перекрест происходит частично в спинном, частично в продолговатом мозге. Посредством восходящих путей афферентная импульсация от рецепторов передается к определенным воспринимающим областям коры. Они здесь заканчиваются преимущественно в четвертом слое в виде сильно разветвленного дерева в зоне распространения звездчатых клеток с околоклеточной сетью, как это показано на рис. 1 и 2. Особенно хорошо это проявляется в процессе онтогенетического развития (рис. 9—I). Эти прямые чувствительные пути принято называть специфическими поскольку они заканчиваются в определенных воспринимающих областях коры. Но все восходящие пути связаны коллатералями с ретикулярным образованием. А это образование имеет прямую связь с корой через таламическую восходящую систему, которая оканчивается в коре более или менее диффуано во всех ее областях, но в основном в ассоциационных. Эту систему афферентной импульсации принято называть неспецифической. Ее окончания встречаются во всех слоях коры, но не в виде метелок, а в виде отдельных ветвящихся волокон (см. рис. 2, с, d). В онтогенезе они развиваются одновременно с апикальными дендритами и в конце концов контактируют с шипиками по всей длине дендритов (рис. 9—II) (М. ШайбельиА. Шайбель, 13). Часть чувствительных путей от органов слуха, зрения и лабиринтов проходит через мозжечок. Здесь они прерываются и последующие нейроны через ретикулярную формацию достигают коры по неспецифической системе (см. рис. 8, В).
Для каждого рецептора имеется своя система передаточных станций. И каждая такая станция включена как в восходящий путь от рецептора до коры, так и в обратный, нисходящий путь. Поэтому каждая ядерная станция связана двойными путями, образуя нервные круги как с вышележащими, так и с нижележащими ядрами, а также с эффекторами и рецепторами. Следовательно, центростремительный путь от рецепторов доходит до коры больших полушарий, а центробежный путь от коры кончается не только в эффекторах, но и в рецепторах (см. рис. 8, А, Б, В, Г) (подробно о центробежных путях будет изложено в гл. IV). Нейроглия в коре большого мозга. Характерным для коры головного мозга является наличие всех основных и переходных форм нейроглии. Кора богата всеми этими клетками и тем самым отличается от других отделов центральной нервной системы. При этом каждый вид нейроглии расположен своеобразно. Шредер (14) и Снесарев (15) дают подробное описание распространения астроцитов в коре головного мозга. Первый поверхностный слой коры представлен волокнистыми астроцитами, в нижней его части появляются протоплазматические и переходные формы астроцитов (имеющие как протоплазматические, так и волокнистые отростки); но во II слое преобладают протоплазматические астроциты; в /// они более маленькие; в IV слое среди протоплазматических астроцитов начинают появляться переходные формы; в V — уже преобладают переходные формы, а в самом нижнем VI слое встречаются только волокнистые астроциты, продолжающиеся непосредственно в белое вещество. Как видно, протоплазматические астроциты преобладают в средних слоях коры, тогда как волокнистые астроциты расположены в поверхностном и глубоких слоях. Эти астроциты специальными ножками связаны со стенкой сосуда (рис. 10). В поверхностном слое расположены волокнистые астроциты, которые своими многочисленными короткими отростками фиксируют мягкую мозговую оболочку и сосуды, а длинными отростками проникают в глубь коры. Создается представление, что расположенные в поверхностном и в VI слое волокнистые астроциты своими отростками создают единую систему эластичных волокон, связанных между собой с помощью кровеносных сосудов, образуя остов для клеточных элементов коры мозга. По подсчету Криспин-Екснера (16), из общего числа глиальных клеток зрительной коры астроглия составляет 61,5%.
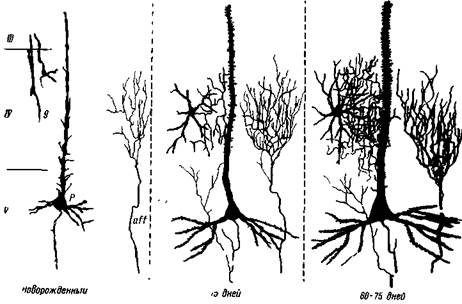
Рис. 9— I. Развитие пирамидных и звездчатых нейронов и окончаний афферентных волокон специфической системы в сенсомоторной коре котенка g — звездчатые клетки (короткоакоошше), которые первоначально представлены в виде биполярных клеток; р — пирамидный нейрон; off — афферентное волокно с небольшим конечным разветвлением. Одновременно с сильным развитием звездчатых клеток сильно нарастает ветвление афферентных окончаний и они достигают полного развития на 60-й день. Пирамидный нейрон также достигает полного развития к этому времени (М. Шайбель и А. Шайбель)
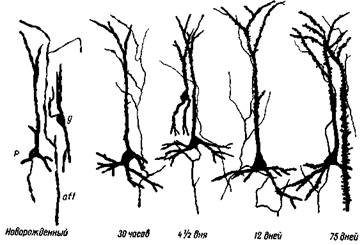
Рис. 9— II. Развитие дендритного аппарата пирамидных нейронов и терминальных окончаний кортикопетальных волокон сенсомоторной коры котят g _ звездчатый нейрон; р — пирамидный нейрон; aff — окончание афферентного волокна неспецифической системы. Последнее с течением времени растет и все лучше и лучше контактирует с шипиками по длине всего дендрита (М. Шайбель и А. Шайбель)
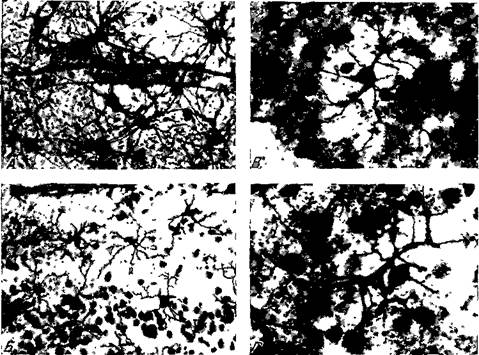
Рис. 10. Глиальные элементы коры головного мозга обезьяны (Macacus rhesus) А — волокнистые астроциты с отростками связаны со стенкой кровеносного сосуда. Б — кора зрительной области — I и II слой. Микроглиоциты расположены характерно' область распространения разветвлений одного микроглиоцита не переходит на территорию разветвления смежных микроглиоцитов. Отростки клеток, в основном в молекулярном слое, снабжены «шипиками» Импрегнация по Мийгава-Александровской. В и Г — V слой зрительной области Микроглиоциты своими отростками приходят в контакт со всеми клеточными элементами, расположенными в области их разветвления (по Микеладзе) Олигодендроглия широко распространена в коре головного мозга и, в отличие от протоплазматических астроцитов, по направлению к белому веществу число олигодендроцитов увеличивается. В первом слое коры они единичны, тогда как во // и /// слоях они сгруппированы по 2—3 клетки, причем некоторые из них выступают как спутники нейронов; в IV и V слоях эти группы состоят примерно уже из 4—5 клеток, и в VI слое их число еще увеличивается. Олигодендроциты составляют 29,0% из общего числа клеток нейроглии (Криспин-Екснер, 16). По старым данным Шпильмейера (17), перинейрональные, так называемые «сателлиты», олигодендроциты расположены главным образом в /// слое. Эти клетки, как замечает Пенфилд (18), существуют в полном симбиозе с нейронами. В последнее время была показана их высокая метаболическая активность. Они принимают участие, между прочим, в водносолевом обмене. Среди глиальных элементов коры мозга в количественном отношении микроглия занимает небольшое место. Микроглиоциты расположены в коре равномерно и притом в шахматном порядке, наподобие протоплазматических астроцитов. По выражению Снесарева (15), «каждый микроглиоцит имеет свой район действия». Область распространения отростков одного микроглиоцита не переходит на территорию смежного микроглиоцита (рис. 10, Б). В последнее время было показано, что каждый микроглиоцит со своими отростками связан с определенными нервными и глиальными клетками и сосудами, расположенными в области их разветвления (рис. 10, В и Г), и как бы объединяет все эти элементы.
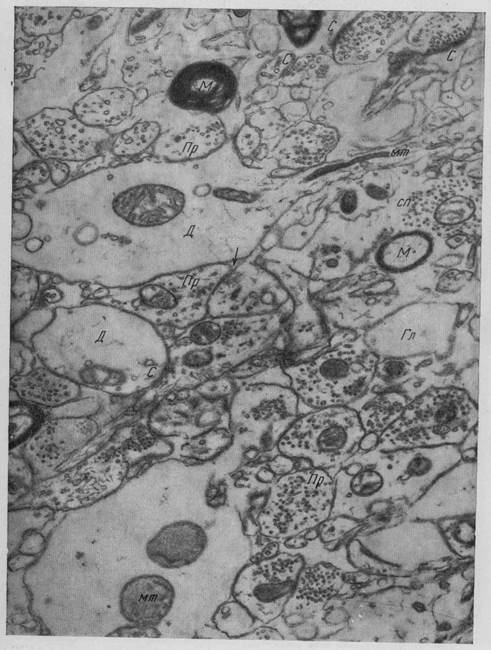
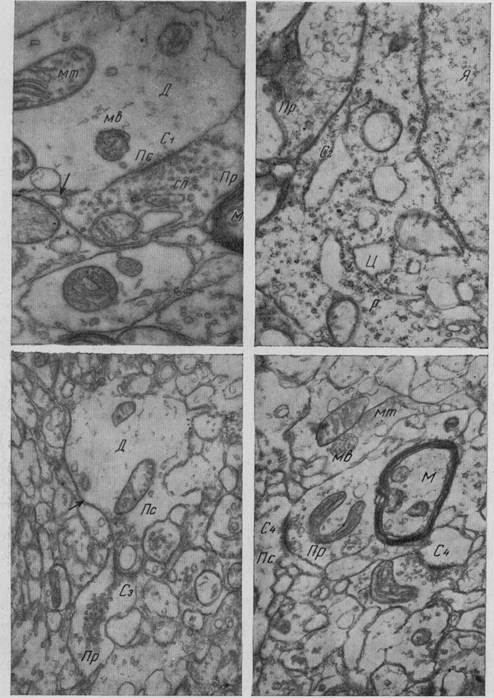
Рис. 11. Кора головного мозга собаки (участок нейропиля) Пр - пресинаптические терминали, Д - дендритные терминали, М - миелиновое волокно мт митохондрия, С — синапс, сп — синаптические пузырьки, Гл — глиальный отросток стрелкой указана межклеточная щель. Ув. 22 000 (Микеладзе)
Рис. 12. Кора головного мозга собаки C1 — синапс II типа на терминали дендрита (Д), мв — мультивезикулярное тело в постсинаптическом участке, С2 — аксо-соматический синапс II типа, С3 — синапс II типа на терминали дендрита, видна инвагинация постсинаптического участка (Пс) в пресинаптический отросток (Пр), постсинаптический отросток содержит крупный пузырек, С4 — синапс II типа образует по два огранитенных синаптических контакта, Я — ядро нейрона, Ц — цистерны эндоплазматической сети, р — рибосомы. Ув. 20 000— 40 000 (Микеладзе)
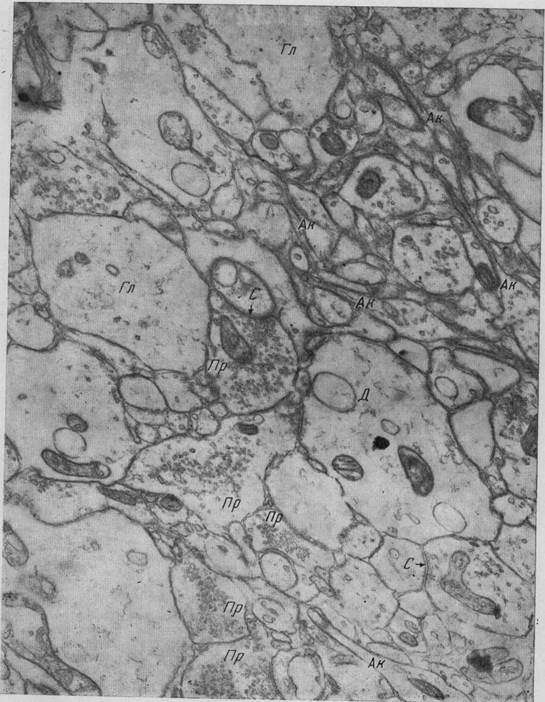
Рис. 13. Молекулярный слой коры головного мозга собаки С — синапс I типа, группа пресинаптических отростков (Пр) находится в непосредственном контакте друг с другом, Ак — тончайшие аксонные терминали, Д — терминаль дендрита, Гг — глиальный отросток. Ув. 25 000 (Микеладзе) Микроглиоциты коры головного мозга характеризуются сравнительно с другими участками серого вещества мозга более сложной структурой (причудливое разветвление отростков, придающее им «мохнатый» вид). Микроглиоциты несколько преобладают в средних слоях коры мозга, а именно в областях, богатых капиллярными сетями. На фронтальных срезах мозга, в области зрительной коры обезьяны, по подсчету Микеладзе (19), Микроглиоциты представлены в / слое в среднем 4,5%, во // — 6,0, в /// — 11,9, в IV— 38,0, в 7 — 30,9 и VI слое —8,7%. Микроглиоциты различных слоев коры головного мозга отличаются друг от друга. Так, микроглиоциты, расположенные в первом слое, сравнительно меньше микроглиоцитов, расположенных в более глубоких слоях, но по степени разветвления не уступают им. Глиальные элементы и их отростки наряду с кровеносными сосудами полностью заполняют межклеточные пространства, т. е. пространства между нейронами и их отростками и нервными волокнами; это хорошо продемонстрировано электронномикроскопически. Расположение глиальных элементов зависит в значительной мере от нейроно- и ангиоархитектоники коры.
|
||||||||
|
|
Последнее изменение этой страницы: 2017-02-21; просмотров: 244; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 13.59.54.188 (0.011 с.) |