Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Онтогенез электрической активности коры мозга животных.Содержание книги
Поиск на нашем сайте Электрическая активность коры больших полушарий головного мозга котят регистрируется в первые же часы после рождения (Петерсен, Перри, Хинвич, 47). У двух-, трехдневных котят электрокортикограмма (ЭКоГ) теменной и затылочной областей состоит из низковольтных нерегулярных ритмов, 4 — 6 в сек. (Шайбель и Шайбель, 48; Джавришвили, 49), перемежающихся с медленными волнами. Амплитуда волн ЭКоГ редко превышает 50 мкв. В сенсомоторной области частота волн ЭКоГ около 10 колебаний в сек. (Шерер и Экономо, 50). К концу первой недели жизни постепенно увеличивается частота. У 6—8дневных котят медленные волны более часты. Между 5—10 днями постнатальной жизни наблюдается возникновение отдельных разрядов веретенообразных волн, имеющих частоту 6—12 в сек., к концу 2 недели ЭКоГ большей частью состоит из прерывистого, а иногда сплошного ряда веретенообразных волн. Через 2—3 дня после возникновения веретенообразных раз-
Рис. 92. ЭКоГ кошки в различные периоды поотнатального развития Расположение отводимых точек показано на схеме мозга и обозначено цифрами А — 1,5-дневного ьотенка, Б—3 дня, В—9 дней, Г— 16 дней, Д — 30 дней, Е — ЭКоГ взрослой кошки (Джавришвили) рядов, обычно следует первое появление альфаподобной активности в затылочной области, что является показателем того, что пришло время открытия век. В дальнейшем, с увеличением возраста животного, ЭКоГ все больше напоминает ЭКоГ взрослой кошки и к 2—3-месячному периоду не отличима от нее (рис. 92). Что касается синхронности разных областей, то у котят до 7—9-дневного возраста волны ЭКоГ синхронны почти по всей поверхности коры. Очевидно, в этом возрасте ЭКоГ всецело обусловливается импульсацией из более развитых подкорковых образований. С возрастом синхронность регулярных волн изменяется. В первую очередь асинхронной относительно других областей коры становится соматосенсорная кора, также в первую очередь изменяется спонтанная активность (по частотной и амплитудной характеристике) в этой области. Возможно, это обусловлено тем, что соматосенсорная кора раньше других начинает созревать и раньше других проекционных областей связывается с внешней средой: раньше других получает афферентную импульсацию. Итак, изучение ЭКоГ в постнатальном развитии свидетельствует о том, что со дня рождения постепенно увеличивается частота и амплитуда волн ЭКоГ, они синхронны во всех частях коры в первые дни, но выделяются доминирующие частоты, и лишь потом наступает асинхронность между разными частями коры. Онтогенез ЭЭГ человека по данным ранних работ (Бергер, 5; Линдсли, 51) и некоторых поздних (Хилл, 52), запись ЭЭГ с поверхности кожи головы новорожденного ребенка не показывает спонтанной электрической активности. Однако другим исследователям удалось регистрировать ЭЭГ не только у новорожденного, но и у плода (Линдсли, 53). У 3—4-месячного плода ЭЭГ представляет собой нерегулярные волны с амплитудой 10—90 мкв с нанизанными на них регулярными волнами (Окомото и Кирике, 54, 55). У 5—6-месячного плода могут возникнуть ритмические волны частотой 8—15 в сек. и амплитудой меньше чем 30 мкв, но обычно вместе с медленными волнами, частота которых 0,5—1 в сек. Эти волны, продолжающиеся до 5 сек.разделяются длительными периодами электрического молчания (Эллингсон, 56). У новорожденных детей при отведении от кожного покрова черепа электрическая активность регистрируется в виде очень нерегулярных колебаний малой амплитуды. Частотный анализ показывает, что ЭЭГ детей с самого первого дня жизни содержит частоты от 0,5 до 30 в сек. На каждой стадии развития анализ активности показал, что набор основных частотных составляющих различен. Однако доминирующие составляющие медленно замещают друг друга с возрастом. У новорожденных детей характерный рисунок ЭЭГ состоит из дельта — активности низкого вольтажа с совершенно нерегулярными частотой и амплитудой и малой синхронизацией между двумя полушариями (Эллингсон, 57). До 3—4-месячного возраста в затылочной области возникает ритм частотой 4 в сек., но в ЭЭГ бодрствующего ребенка наблюдается мало изменений. Главные изменения наступают к концу года, когда частота ритма быстро увеличивается, почти удваивается и достигает частоты альфа-ритма — 8 в сек. Увеличивается также его амплитуда. Затылочный альфа-ритм постепенно учащается, достигая к 12—13 годам частоты альфа-ритма взрослых (Брейзье, 40). У новорожденных детей потенциалы мозга при сенсорных раздражениях регистрируются в виде неспецифических и первичных ответов. Неспецифический ответ у новорожденных очень своеобразен и характеризуется тем, что при периферическом раздражении возникают генерализованные, полиморфные разряды медленных волн по всей коре. Эти ответы легче получаются во время сна. Проприоцептивное раздражение более эффективно в вызывании такого ответа, слуховое менее эффективно, а зрительное раздражение, которое редко вызывает этот ответ, еще менее эффективно. Вызванный ответ на световое раздражение у некоторых детей состоит только из начальной положительной фазы, у других за положительной фазой следует отрицательная. У некоторых детей первичный ответ состоит только из отрицательной фазы. Отсутствие положительной фазы является показателем незрелости коры мозга (Эллингсон, 57). Первичные ответы детей на световое раздражение отличаются от первичных ответов взрослого человека «утомляемостью» и большим латентным периодом (около 190 мсек.). Латентность с возрастом уменьшается и к 3-месячному возрасту достигает 100 мсек. Уменьшение латентного периода связывают с увеличением скорости проведения в афферентных волокнах (Эллингсон, 57). Все вышеперечисленные изменения в ЭЭГ и первичных ответах отражают степень созревания головного мозга. В течение пренатального и постнатального периодов онтогенеза поле ветвления дендритов и аксонов нейронов, которые устанавливают контакты между ними, значительно увеличивается. Особенно интенсивно в период пренатального онтогенеза развиваются большие пирамиды V слоя и звездчатые клетки, которые находятся в четвертом и пятом слоях. Эти системы нейронов, как уже рассматривалось нами в I главе, развиваются и созревают в онтогенезе раньше всех, будучи в основном сформированы при рождении (Поляков, 58). Изменения ЭЭГ во время сна. Во время нормального сна у животных (кошек) колебания бета-ритма временами выпадают, в это время возникают длительные нотенциалы частотой 2—3 в сек. — дельта-волны (рис. 93)* Пробуждение животного вызывает исчезновение длительных и появление быстрых потенциалов (Рейнбергер и Джаспер, 59; Коган, 34; Цкипуридзе, 14). Примерно то же наблюдается на препарате при засыпании (что также проявляется в сужении зрачков) — в коре кошки возникают медленные потенциалы с ритмом 2—4 в сек. (Бремер, 60). Более того, эти же изменения альфа- и бета-волн наблюдаются при засыпании и пробуждении, если кора отделена от всех подкорковых образований путем перерезки объединяющих их путей. При засыпании животного исчезают быстрые потенциалы и возникают медленные — по ритму 7—8 в сек. При пробуждении наблюдаются обратные явления (Хананашвили, 126). Но у кошек, как теперь установлено^ периоды сна с наличием в коре медленных потенциалов чередуются с периодами глубокого сна, когда в коре регистрируются быстрые колебания малой амплитуды (Демент, 61). У человека ЭЭГ во время сна меняется очень своеобразно и закономерно. Выделяют 5 стадий развития ЭЭГ (Лумис, Гарвей и Хобарт, 62). Первоначально теменно-затылочный альфа-ритм несколько усиливается; с наступлением дремоты амплитуда альфа-волн прогрессивно уменьшается, они начинают наступать более короткими группами и, наконец, альфа-ритм исчезает; часто при этом наступает период полного затишья в ЭЭГ (Бакурадзе и Нарикашвили, 63) (рис. 94) и эта стадия обозначается как стадия А, а период исчезновения альфа-ритма предложено обозначать «а» малым (Кратин, 26). В стадии А, пока существует альфа-ритм, внешние раздражения вызывают его депрессию. Нерегулярные медленные колебания основной электрической активности начинают усиливаться еще в дремотном состоянии. С развитием сна ониеще более усиливаются, особенно в центральных и лобных областях; появляются нерегулярные уплощенные тета-волны. Эта стадия обозначается как
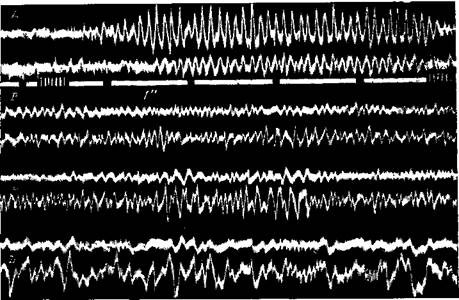
Рис. 93. Кошка с хроническими отводящими электродами над соматосенсорной (/) и слуховой (2) областями коры, в сстевидном образовании среднего иозга (3) и во внутренней коленчатой теле (4) Запись на чернильнопишущем электрсэнцефалографе. А — кошка в бодрствующем состоянии; Б — кошка спит (Ройтбак и Бутхузи)
Рис. 94. Электрическая активность коры большого мозга человека во время сна Запись шлсйфным осциллографом. В каждой записи верхняя кривая — от затылочно-теменных долей, нижняя — от центрально-лобных. А — перед сном; до закрытия глаз слабые нерегулярные колебания основной электрической активности: с закрытием появляется альфа-ритм: сильно в теменно-затылочной области, слабо — в центрально-лобной. В — дремотное состояние через 20 мин. после разрешения спать; в обоих участках основная активность усилена, моментами ровный ритм 18—22 в сек., альфа-волны ослаблены. С — через 30 мин. от начала засыпания, спит, на зов не реагирует. Основная электрическая активность еще больше усилена, особенно в лобной области, местами ровный ритм 14— 16—22 в сек, альфа-волны отсутствуют. D — через 1 час 45 мин. храпит, альфа-волны отсутствуют; основная электрическая активность еще более усилена. Временами наступают дельта-подобные волны длительностью в 0,25—0,4 сек., на которые наложены более слабые потенциалы короткой продолжительности в 0,1—0,05 сек. Эти волны значительно сильнее в лобной области, чем в затылочно-теменной (Бакурадзе и Нарикашвили) стадия В. В стадии В периферические раздражения часто вызывают вспышки альфа-ритма — т. е. как бы переход в стадию А. С дальнейшим углублением сна появляется так называемый ритм «веретен» 14—18 в сек., перемежающийся с тета- и дельта-волнами (стадия С). Во время глубокого сна доминируют дельта-волны (стадия D). Они наступают то одиночками, то группами, и притом более или менее синхронно в разных участках коры; но синхронность дельта-волн, как и колебаний основной активности, во время сна выражена слабее, чем у колебаний основной активности и у альфа-волн в бодрствующем состоянии (Девис и сотр., 64; Бакурадзе и Нарикашвили, 63). Под влиянием звуковых раздражений дельта-волны в некоторых случаях изменяются: они ослабевают или устраняются, т. е. звуки (шум) действуют на дельта-волны во время сна как на альфа-волны в бодрствующем состоянии. В стадии еще более глубокого сна (стадия Е) в ЭЭГ доминируют нерегулярные уплощенные колебания. Внешние раздражения умеренной интенсивности не вызывают изменений ЭЭГ. В этой стадии периодически возникают в определенных областях коры сильные двуфазные колебания, так называемые судорожные разряды (Бакурадзе и Нарикашвили, 63); видимо, они возникают при судорожных вздрагиваниях спящего. Такой период сна сейчас называют парадоксальным сном (подробно об этом сне см. в след, главе). Сходные стадии изменения электрической активности коры при развитии сна установлены и у кошек (Кратин, 26).
|
||
|
|
Последнее изменение этой страницы: 2017-02-21; просмотров: 338; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.147 (0.012 с.) |