Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Возбуждение двигательных участков.Содержание книги
Поиск на нашем сайте
Процессы возбуждения в коре большого мозга до последнего времени изучались по тем биоэлектрическим токам возбуждения, которые регистрировали в мышцах при корковых двигательных реакциях. Этим путем было установлено, что возбуждающий импульс в коре большого мозга продолжается такое же короткое время, как и
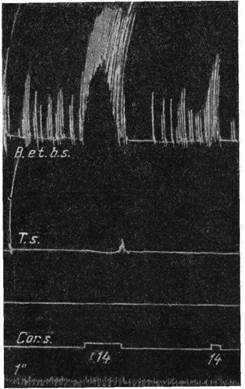
Рис. 77. Явление суммации и облегчения при раздражении коры Обезьяна Macacus rhesus. Верхняя кривая — от сгибателя плеча m. supinator longus, а нижняя — от разгибателя m. triceps br. Раздражается двигательный участок в противоположном полушарии. Всего 6 тетанических раздражений продолжительностью около 1 сек. с интервалами около 4 сек. Первое раздражение было субминимально: оно не дало эффекта, второе дало ничтожный эффект — слабое сокращение на сгибателе, а все последующие раздражения произвели прогрессивно нарастающие эффекты (Грэим Броун) в центральной нервной системе вообще. Корковое возбуждение, безусловно, также сопровождается абсолютной рефракторной фазой и следует одно за другим ритмически. При ритмическом раздражении коры регистрируемые в мышце биопотенциалы меняются в зависимости от частоты и силы раздражения коры совершенно так же, как было установлено для спинномозговых рефлексов (Гофман, 73; Купер и Денни-Броун, 74). Это, конечно, было бы невозможно, если бы возбуждение коры не протекало ритмически или если бы кора мозга не была способна к высокому ритму возбуждения.
Ритмическое течение возбуждения в двигательных участках подтверждается и при изучении электрических явлений непосредственно в них самих. Раздражение двигательных участков или их спонтанное возбуждение всегда производит ритмическое колебание потенциала очень изменчивой частоты, до 100—200 в сек. и выше. Эти электрические явления должны выражать деятельность как раз тех нервных кругов, которые при этом возбуждаются в области двигательных участков коры. Наряду с быстрыми колебаниями потенциала в двигательных участках возникают медленные колебания, которые должны представлять собой результат суммации множества разновременно возникающих дендритных биотоков (Беритов и Гедеванишвили, 75). При одновременном изучении биотоков двигательной зоны коры и движений конечностей у животных оказалось, что возникающие в коре медленные биотоки в ритме 20—40 в сек. находят свое выражение в ритме клонических подергиваний соответствующей конечности. При параллельной же регистрации биотоков двигательного участка коры и соответствующей мышцы было установлено, что означенные электрические колебания в коре в ритме 20—40 в сек. более или менее совпадают с группировкой быстрых колебаний соответствующей мышцы. Именно, каждому медленному колебанию потенциала в коре отвечает одна группа быстрых колебаний в мышце (Эдриан, 76) (рис. 78). Вследствие этого ритмическое течение биотоков в мышце принимает двоякий характер: с одной стороны, мышечные биотоки в высоком ритме 150— 200 в сек; а с другой,— их группировка в ритме 20—40 в сек. Ритм группировок мышечных биотоков отвечает ритму медленных колебаний в коре дендритного происхождения, а ритм самих мышечных биотоков 150 —200 в сек., должно быть, обусловливается возбуждением пирамидных клеток импульсами возбуждения из нервных кругов. Как подробно будет изложено в следующей главе, нервные круги в коре большого мозга возбуждаются не беспрерывно, а периодически. Это обусловливает как периодическое активирование дендритов в ритме 20—40 в сек., так и периодические разряды импульсов, которые через пирамидные и экстрапирамидные пути производят периодическое возбуждение скелетных мышц.
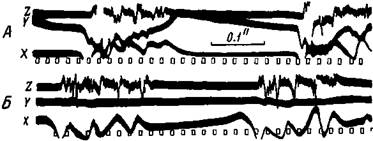
Рис. 78. Биотоки возбуждения коры и мышцы после электрического раздражения двигательного участка передней конечности обезьяны А— электрограммы: X — при отведении ближайшего, У — несколько более удаленного участка коры, Z — при отведении мышц передней конечности, Б — электрограммы в таком же опыте X — при отведении ближайшего участка, Y — при отведении отдаленного участка задней конечности, Z — от мышц передней конечности (Эдриан) Аналогичные результаты получали Эдриан иМоруцци (77) при одновременном изучении биотоков двигательных участков коры и нисходящих пирамидных путей в продолговатом мозге животного. Между этими токами существует большая согласованность, но и в этом случае очень часто одной волне коркового потенциала соответствует несколько быстро протекающих импульсов в пирамидных путях. Иной раз после воздействия на кору некоторых ядов, таких как стрихнин, в пирамидных путях регистрируются очень высокие ритмы — до 500—1000 в сек., но небольшими группами: по 10—80 колебаний. Приведенные выше наблюдения указывают, что двигательные участки коры большого мозга, или, точнее, нервные круги их, разряжаются импульсами в таком высоком ритме, какой вообще свойственен нервному волокну. Но ввиду периодической деятельности нервных кругов, как активирование дендритов коры мозга, так и возбуждение пирамидных и экстрапирамидных путей происходят периодически в ритме 10—40 в сек. Многие авторы, изучавшие произвольную иннервацию скелетных мышц, наблюдали с помощью струнного гальванометра, что ритм 40—50 в сек. доминирует над другими более редкими и частыми ритмами. Они считали этот ритм за основной ритм возбуждения центральных элементов. Более высокий ритм объясняли разновременной деятельностью центральных элементов (Эдриан и Бронк, 78; Правдич-Неминский, 79; Рихтер, 80). Предполагалось, что возникновение импульсов возбуждения в значительном числе центральных элементов совпадает во времени и этим самым обусловливается низкий ритм 40—50 в сек; некоторое же число их возбуждается в разное время и ве- дет к возникновению мелких волн высокого ритма (Эдриан). И действительно, при регистрации биотоков небольшого пучка мышечных волокон одной двигательной единицы, иннервируемой одним двигательным нервным волокном, ритм возбуждения меняется всего от 5 до 50 в сек. (Ваххольдер, 53; Рихтер, 80; Линдсли, 81). При самых слабых сокращениях двигательной единицы этот ритм составляет 5—10 в сек. С усилением произвольного сокращения ритм возбуждения повышается, но не превышает 50 в сек. Наблюдаемое при этом усиление сокращения всей мышцы зависит не только от учащения возбуждений двигательной единицы, но в значительной мере и от увеличения числа активных двигательных единиц (Линдсли, 81). Очевидно, более высокий ритм, 150—200 в сек., регистрируемый от больших участков мышцы или целой мышцы, обусловливается разновременным течением возбуждения во многих двигательных единицах. Отсюда следует, что нормальный ритм возбуждения корковых двигательных элементов при произвольной иннервации очень низок: 5—50 в сек.
Этот низкий ритм, конечно, нельзя считать выражением лабильности корковых двигательных элементов, ибо, как указывалось выше, корковые элементы могут отвечать и более высоким ритмом возбуждения, вдвое — втрое превосходящим нормальный. Как указывалось выше, при отведении изолированных пирамидных и особенно звездчатых нервных клеток коры ритм возбуждения оказывается очень изменчивым, но доходящим в некоторых случаях до 600 в сек. (Джаспер и сотр., 82). Очевидно, низкий ритм, до 50 в сек., является наиболее благоприятным ритмом возбуждения корковых двигательных элементов — таким ритмом, при котором они длительно могут работать без утомления. И действительно Линдсли (81) показал, что при длительных произвольных сокращениях, до 15—30 в мин., нормальный ритм корковой иннервации не меняется, значит, не происходит утомления двигательных элементов коры. Меняется только амплитуда токов возбуждения мышцы, что, должно быть, зависит от утомления менее лабильных координирующих элементов спинного мозга или самой мышцы.
|
|||||||||||||
|
|
Последнее изменение этой страницы: 2017-02-21; просмотров: 251; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.139.86.74 (0.01 с.) |