Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Сенсорная функция звездчатых нейронов первого типа.Содержание книги
Поиск на нашем сайте Есть основание утверждать, что субъективное отражение внешнего мира (ощущения, восприятия, представления, образы) возникает в первичной зоне анализаторов под влиянием специфической афферентной импульсации. Это следует из того, что по наблюдениям на людях появление вызванных потенциалов, регистрируемых в первичной зоне анализаторов, и субъективные ощущения тех раздражений, которые вызывают эти потенциалы, совпадают. Когда в результате отвлечения внимания или при усиленной умственной работе (например, решении арифметической задачи) в коре и восходящих таламических путях ослабевают вызванные потенциалы в ответ на определенные периферические раздражения, тогда ослабевают и субъективные переживания от этих раздражений (Эрнандец-Пеон и др., 13). Особенно демонстративно выявляется параллелизм между электрическими потенциалами и субъективными ощущениями при сопоставлении пиковых потенциалов, регистрируемых от отдельных нейронов первичной зоны зрительного анализатора у кошки, с одной стороны, и зрительными ощущениями человека,— с другой. Было найдено двенадцать разного рода параллельных явлений. Например, при усилении светового раздражения интенсивность светового ощущения нарастает по известному закону ВеберФехнера. Соответственно и частота разрядов определенных нейронов зрительной коры увеличивается приблизительно в логарифмической зависимости от величины световой интенсивности, если световая адаптация сохраняется постоянной. Субъективное слияние мелькающего света у человека происходит при частоте около 50 в сек., а предельный ритм возбуждения зри-
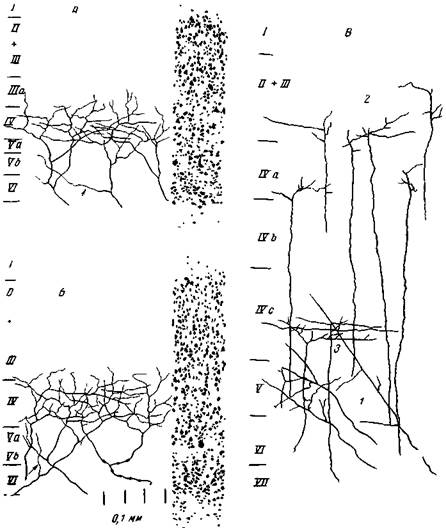
Рис. 20. Звездчатые нейроны «Слева — по Полякову (микрофотография) и оправа — по Чангу (полусхематический рис). На обоих рисунках ясно выступает четковидное строение дендритов
Рис. 21. Синапсы А — на звездчатой клетке из IV слоя 18 поля. Б — на большой звездчатой клетке с длинными нисходящими аксонами из IV слоя 17 поля. Синапсы в виде колец и петелек видны как на теле, так и на дендритах (Энтин)
Рис. 22. Распределение специфических афферентных таламических волокон в зрительном анализаторе А и Б — в разных участках area striata кошки С правой стороны дается относите гьная ширина корковых слоев; слева — таламические волокна, стрелки указывают на тонкие коллатерали, которые отходят в пятом слое (О'Лири) В — афферентные волокна в 17 поле церкопитека. Волокна оканчиваются как в 1Услое, так и в слоях У и III (Школьник-Яррос) тельных нейронов при мелькающем свете — около 50 в сек. В особых случаях при повышении интенсивности света нарастает как субъективный ритм мелькания, так и ритм объективного разряда зрительных нейронов (Юнг, 14). Далее, из наблюдений на людях во время хирургических операций установлено, что обычно при электрическом раздражении коры субъективныз ощущения возникают только при раздражении воспринимающих областей. При этом, в зависимости от раздражаемой первичной зоны, больной ощущает свет, цвет, звук, прикосновение, дрожание пальцев и т. д. Но эти ощущения не отражают конкретных внешних явлений (Пенфилд, 15). Это и понятно, ибо возникновение образов предполагает участие определенного очень сложного комплекса корковых нейронов, пространственно расположенных в разных участках коры, одновременное возбуждение которых не может возникнуть при электрическом раздражении коры. Мы предполагаем, что паукообразные и им подобные клетки с околоклеточной сетью в IV слое первичных зон являются теми нервными элементами кори, которые вызывают при своем возбуждении субъективные переживания. Поэтому звездчатые клетки с околоклеточной нервной сетью можно назвать сенсорными. Наличие же у этих звездчатых нейронов контактов своего аксонного разветвления на собственных дендритах и даже на собственном теле, как указывалось выше, создает возможность самовозбуждения. Поэтому каждый раз в ответ на один афферентный импульс постсинаптическое возбуждение клетки должно значительно усиливаться и удлиняться в результате повторного возбуждения аксона. Очевидно, только при такой активации звездчатых клеток возникает субъективное переживание воспринятого объекта. Как известно, тот высокий уровень возбудимости звездчатых нейронов, от которого зависит восприятие внешних объектов и их проецирование во внешней среде, т. е. субъективное отражение внешнего мира, обусловливается действием определенной части неспецифической системы. При ее искусственном выключении путем перерезки или при физиологическом выключении путем торможения животное перестает воспринимать внешний мир. Из этого следует, что готовность или способность сенсорных звездчатых нейронов к активации в ответ на внешние раздражения существенно зависит от ретикулярной системы. Ее восходящие импульсы, действуя на эти нейроны, обусловливают в них тот уровень возбудимости или чувствительности, который необходим для восприятия внешнего мира. Сенсорное свойство звездчатых клеток IV слоя ни в коем случае нельзя считать функцией внешнего строения нейрона, т. е. особенностей разветвления его аксонов и дендритов в виде образования околоклеточной сети. Безусловно, внешняя форма сенсорных нейронов может быть самая различная. Однако паутинообразная форма сенсорных звездчатых клеток без аксонных связей с другими клетками, вероятно, наиболее характерна для высших животных и человека. Это внешнее строение приспособлено к наилучшему способу повторной максимальной активации клетки под влиянием действующих на нее афферентных импульсов. Поэтому такие звездчатые и даже паутинообразные клетки могут существовать и в других отделах мозга, а не только в коре. Производство дифференцированных субъективных ощущений является филогенетически выработанным свойством нервной цитоплазмы. Только благодаря такому свойству цитоплазмы определенные нервные клетки стали основой психической деятельности, ибо на субъективных ощущениях зиждется восприятие внешнего мира и создание образов или конкретных представлений о внешнем мире. То обстоятельство, что лишь некоторые ближайшие звездчатые паутинообразные клетки связаны между собой, образуя гнезда, а между этими гнездами нет непосредственных связей, имеет глубокий биологический смысл Звездчатые нейроны TV слоя должны воспринимать и отражать внешний мир; они должны приходить в активное состояние одновременно под влиянием раздражителей внешнего мира, чтобы создать конкретные его образы. Иррадиация возбуждения в системе звездчатых нейронов могла бы только помешать такому образному восприятию внешнего мира и быстрой смене образов в связи с изменчивостью внешних воздействий. Тот факт, что значительная часть паутинообразных звездчатых клеток не имеет также аксонных связей и с пирамидными клетками, указывает, что не все сенсорные звездчатые клетки участвуют в прямой передаче возбуждения к пирамидным клеткам для осуществления хотя бы ориентировочного движения головы. Их основная функция — производить субъективное отражение внешнего мира, создание его образов. Однако нельзя утверждать, что звездчатые клетки с околоклеточной сетью не играют никакой роли в вызове внешней реакции. Когда эти клетки возбуждаются при репродукции образа какого-либо объекта, то возникшие в них медленные потенциалы могут влиять электротонически на другие нервные элементы, прежде всего, на афферентные таламические волокна, которые посредством своих коллатералей оканчиваются как на звездчатых нейронах,
так и на пусковых клетках ориентировочной реакции. При репродукции образа под влиянием ассоциационных импульсов в сенсорных звездчатых нейронах возникают медленные постсинаптические потенциалы, электротонически распространяющиеся по дендритам в клетку, а затем выходят через синаптические участки по оканчивающимся на клетке афферентным волокнам.
Здесь мы, должно быть, имеем такое же явление, как в спинном мозге, когда медленные потенциалы двигательных и промежуточных нейронов электротонически выносятся наружу по оканчивающимся на них заднекорешковым волокнам (Беритов, 17). Электротонические токи из звездчатых нейронов по коллатералям афферентных таламических волокон достигают пусковых клеток ориентировочной реакции, например, звездчатых клеток Нахала и пирамидных клеток Мейнерта.А поскольку при выходе тока из этих клеток последние должны деполяризоваться, особенно в области аксонного холмика, то тем самым пусковые клетки приходят в состояние повышенной возбудимости. Ввиду короткого расстояния между пусковыми ориентировочными клетками и звездчатыми нейронами это деполиризующее действие должно быть значительным. Поэтому пусковые клетки должны обладать в этот момент высокой готовностью к реакции на афферентные импульсы, а также на всякие доходящие до них интрацентральные импульсы. Должно быть, здесь мы имеем явление такого же рода, как и при вызове тонических рефлексов: пассивный или активный поворот головы благодаря раздражению шейных и лабиринтных рецепторов лишь сильно повышает возбудимость в определенных нервных центрах. Возбуждение же последних — вызов соответствующего тонического рефлекса, зависит от иных внешних и внутренних раздражений. Рецепторная, сенсорная функция звездчатых клеток ///— IV слоев хорошо выступает при сравнении гистологического строения, во-первых, первичной зоны у низших и высших представителей млекопитающих и, во-вторых, первичной ядерной зоны и периферических отделов, т. е. вторичной зоны, в одной и той же воспринимающей области. Из работы Школьник-Яррос хорошо видно, что у ежа — животного со слаборазвитым зрением в 17 поле зрительного анализатора IV слой развит слабо и очень беден звездчатыми клетками (рис. 23); у собаки, отличающейся лучшим зрением, IV слой развит сильнее и содержит значительно больше звездчатых клеток (рис. 24); у низших обезьян — животных с сильно развитыми зрительными ориентировочно-исследовательскими реакциями IV слой 17 поля еще более широк: он распадается на три густоклеточных слоя, содержащих очень большое количество разнообразных звездчатых клеток (рис. 25). При сравнении централь-
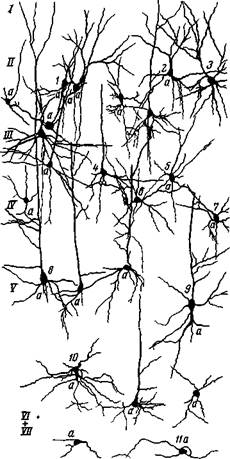
Рис. 24. Нейроны 17 поля зрительного анализатора собаки 1 — клетка I слоя; *, в — пирамидные клетки III слоя, 3,4 — звездчатые клетки; S, 9 — пирамидные клетки IV слоя; 7, ю — звездчатые клетки IV слоя с нисходящим аксоном; 11— крупная пирамидная ь тетка V слоя (Школьник-Яррос) ного 1? поля с периферическими полями 18 и 19, которые прилегают к 17 полю и также принадлежат зрительной воспринимающей области, ясно выступает характерное различие в составе клеток. В полях 18 и 19 звездчатых клеток значительно меньше и они расположены узкой полосой, составляющей IV слой (рис. 26). Весьма вероятно, что и эти клетки имеют отношение к зрительному восприятию. Как известно, у животных после разрушения первичной зрительной зоны зрительная рецепция сохраняется в виде ощущения света и тени. Здесь во II, III и IV слоях господствуют пирамиды и совсем отсутствуют характерные для 17 поля крупные клетки Кахала и мелкие их разновидности, которые, по нашему мнению, служат для вызова ориентировочных движений глаз. В IV слое ядерной зоны кожно-кинестетического анализатора, занимающего у человека и приматов постцентральную область, больше половины всего клеточного состава приходится на звездчатые клетки (Поляков, 18). По Р. Кахалу, такое преобладание звездчатых клеток в IV слое есть анатомическое выражение утонченной функции головного мозга человека. По Лоренте де Но (4), это представление является почти фактическим положе-
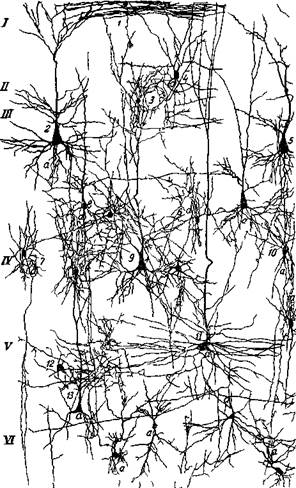
Рис. 25. Нейроны 17 поля зрительного анализатора обезьяны (церкопитека) 2 — пирамидная клетка II слоя, 4, 5, в — пирамидные клетки III слоя, 9, 12 — звездчатые клетки Нахала подслоя IV6; 7,8 — звездчатые клетки подслоя IVa; 11, IS, n, 18— мелкие звездчатые клетки подслоя IVe, IK, 20, 24— звездчатые клетки подслоя IVc; 19 — пирамидная клетка подслоя IVc; 21, 26— крупные пирамидные клетки TV и V с'юев (Школьник-Яррос; нием. По Полякову и Саркисову (1), в этом преобладании нейронов с короткими аксонами и заключается главное отличие коры мозга человека от коры мозга животных в смысле большей сложности и совершенства ее организации. Характерно, что Р. Кахал в книге, посвященной структуре архипалеокортекса (19), приводит типичные звездчатые клетки с околоклеточной аксонной сетью только в отношении обонятельной коры. На них оканчиваются обонятельные волокна. Эти клетки, несомненно, являются сенсорными, производящими запаховые ощущения. По всему видно, что звездчатые клетки IV слоя коры, и прежде всего те из них, которые окружены сетью аксонных разветвлений, имеют прямое отношение к самой высшей функции коры мозга, к субъективному восприятию внешнего мира. Можно утверждать, что основным свойством данных звездчатых клеток является производство дифференцированных ощущений при каждом возбуждении их. Из этих ощущений создаются образы воспринятых предметов, проецируемые во внешней среде (Бериташвилп, 20) (рис. 25).
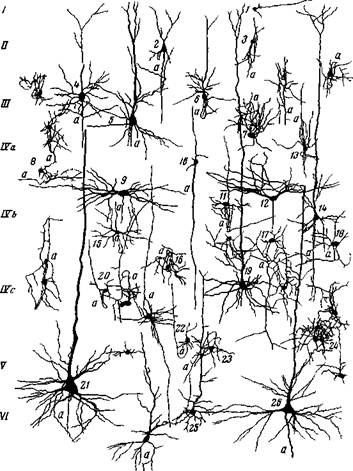
Рис. 26. Нейроны 19 поля зрительного анализатора обезьяны (церкопитека) 1,3,4,6,8 — пирамидные клетки II и III слоев, s,7 — кустовидные клетки III слоя 9, ю — звездчатые клетки IV слоя, и — пирамидная клетка V слоя (Школьник-Яррос) Характерно также, что у человека и высших млекопитающих в коре мышечной воспринимающей области, сенсорная функция которой не связана в значительной мере с ясно дифференцированными субъективными переживаниями, четвертый слой, типичный для других сенсорных областей, отсутствуег. Соответственно звездчатых клеток в двигательной коре очень мало, и они рассеяны по всем слоям коры (Жукова, 21). Этот факт также свидетельствует о том, что в коре восприятие внешнего мира должно быть связано со звездчатыми клетками IV слоя. Итак, звездчатые клетки с паутинообразным разветвлением их аксонов вокруг тела клетки в области дендритов, а может быть и другие звездчатчые клетки, расположенные главным образом в первичных зонах воспринимающих областей, как по физиологическим и клиническим наблюдениям, так и по их количественному нарастанию в связи с филогенетическим развитием воспринимающей функции, являются главными воспринимающими сенсорными элементами коры. Звездчатые клетки, принадлежащие ко второму типу, аксоны которых ветвятся вдали от клетки в вертикальном или горизонтальном направлении, не должны выполнять рецепторной функции. Они, должно быть, играют ту
Рис. 27. Ассоциационные нейроны в коре больших полушарий по Чангу Слева представлены клетки ассоциационных пирамидных нейронов, которые расположены в IV— VI слоях.справа — их окончания в трех верхних слоях же роль передатчика возбуждений, что и мелкие пирамидные клетки с короткими аксонами.
|
||||
|
|
Последнее изменение этой страницы: 2017-02-21; просмотров: 359; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.220 (0.008 с.) |