Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Взаимодействие аллельных геновСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
При полном доминировании один ген полностью подавляет проявление другого гена (выполняются законы Менделя); при этом гомо- и гетерозиготы фенотипически неотличимы. Например, ген желтого цвета семян гороха полностью подавляет ген зеленой окраски, а ген карего цвета глаз у человека — ген голубого их цвета. При неполном доминировании (промежуточном наследовании) доминантный ген не полностью подавляет проявление рецессивного гена. У гибридов первого поколения наблюдается промежуточное наследование, а во втором поколении расщепление по фенотипу и генотипу одинаково — 1:2:1. Например, если скрестить растения душистого горошка с красными и белыми цветками, первое поколение будет иметь розовые цветки:
При скрещивании гибридов первого поколения (с розовыми цветками) во втором поколении получим соотношение по фенотипу и генотипу 1:2:1. Доминантные гомозиготы (АА) будут иметь красную окраску цветков, гетерозиготы (Аа) — розовую, а рецессивные гомозиготы (аа) — белую. Такое явление можно объяснить дозой гена. Доминантный ген (А) детерминирует синтез красного пигмента, при наличии его рецессивной аллели (а) пигмент не образуется (гомозиготы аа — белые). У доминантных гомозигот (АА) два активных гена детерминируют синтез большего количества пигмента, и растения имеют ярко окрашенные красные цветки. Гетерозиготы содержат только один активный ген (А), у них вырабатывается вдвое меньше пигмента, чем у доминантных гомозигот, и окраска их цветков будет бледно-красная (розовая). При сверхдоминировании доминантый ген в гетерозиготном состоянии проявляет себя сильнее, чем в гомозиготном. У мухи дрозофилы имеется рецессивный летальный ген (а) — гомозиготы (аа) погибают. Мухи, гомозиготные по гену А (АА), имеют нормальную жизнеспособность, а гетерозиготы (Аа) живут дольше и более плодовиты, чем доминантные гомозиготы. Такое явление можно объяснить взаимодействием продуктов генной активности. При кодоминировании гены одной аллельной пары равнозначны, ни один из них не подавляет действия другого; находясь в генотипе, оба проявляют свое действие. Типичным примером кодоминирования является наследование групп крови человека по системам АВ0 (группа АВ) и MN (группа MN). Четыре группы крови человека по АВ0-системе определяются наследованием трех аллелей одного гена: I0, IА и IВ (пример множественного аллелизма, см. ниже). При этом 1(0) группа крови обусловлена рецессивным геном I0, П(А) — геном IА, III(Б) — геном IB, a IV(AB) — генами IА и IВ одновременно. Рецессивный ген I0 не детерминирует синтез специфических белков (антигенов) в эритроцитах. Ген IА доминантен по отношению к гену I0 и детерминирует синтез в эритроцитах антигена А. Ген IВ доминантен по отношению к гену I0 и детерминирует синтез в эритроцитах антигена В. Одновременное присутствие в эритроцитах генов IА и IВ обусловливает наличие в них антигенов А и В (IV группа крови). Таким образом, гены IА и IВ не подавляют действия друг друга. Они являются равноценными — кодоминантными. Кодоминирование имеет место и при наследовании у человека групп крови по системе MN. Эта система обусловлена наличием двух аллелей — LM и LN. Ген LM обусловливает наличие в эритроцитах человека антигена М (группа крови М), а ген LN — антигена N (группа крови N). Одновременное присутствие в генотипе обеих аллелей обусловливает наличие в эритроцитах обоих антигенов M и N (группа крови MN). Своеобразные внутриаллельные взаимодействия наблюдаются в случае множественных аллелей. Множественными аллелями называются аллели, которые представлены в популяции более чем двумя аллельными состояниями. Они возникают в результате многократного мутирования одного и того же локуса (участка) хромосомы. При этом помимо доминантного и рецессивного генов появляются еще и промежуточные аллели, которые по отношению к доминантному ведут себя как рецессивные, а по отношению к рецессивному — как доминантные. Так, например, у кроликов. сплошная темная - окраска шерсти обусловлена доминантным геном А, животные с белой окраской шерсти — гомозиготы рецессивные (аа). Сплошная серая окраска (шиншилловая) проявляется у гомозиготных организмов по гену ach, а гималайская (основная масть белая, а кончики ушей, лап, хвоста и носа темные) — у гомозигот ah. Ген А доминантен по отношению ко всем аллелям, ген ach рецессивен по отношению к гену А, но доминантен по отношению к генам а'1 и а; ген ah рецессивен по отношению к генам А и ach, но доминантен по отношению к гену а. Кратко это можно записать следующим образом: A>a ch >a h >a. К разновидностям внутриаллельного взаимодействия генов относится и аллельное исключение, когда у гетерозиготного организма в одних клетках активна одна аллель, а в других — другая. Например, у человека и млекопитающих каждая плазматическая клетка синтезирует только одну (свою) цепь иммуноглобулинов (антител). Другим примером аллельного исключения является инактивация одной из двух Х -хромосом у женского организма (см. гл. 6). Случайный характер инактивации приводит к выключению функционирования в одних клетках материнской Z-хромосомы, а в других - отцовской.
Взаимодействие генов Отклонение от законов Менделя вызывают различные виды взаимодействия генов (за исключением полного доминирования), обусловленного геномным уровнем организации наследственного материала. Различают взаимодействия аллельных и неаллельных генов. Взаимодействие генов одной аллели называется внутриаллельным. Выделяют следующие его виды: полное доминирование, неполное доминирование, сверхдоминирование, кодоминирование и аллельное исключение.
Взаимодействие генов разных аллелей называется межаллельным. Различают следующие его виды: комплементарность, эпистаз, полимерию и "эффект положения". При комплементарности присутствие в одном генотипе двух доминантных (рецессивных) генов из разных аллельных пар приводит к появлению нового варианта признака. Различают три разновидности комплементарного взаимодействия генов. I. Два доминантных неаллельных гена по отдельности не имеют фенотипического проявления, а дополняя друг друга, обусловливают новый вариант признака. Развитие слуха у человека. Для нормального слуха в генотипе человека должны присутствовать доминантные гены из разных аллельных пар - D и Е. Ген D отвечает за нормальное развитие улитки, а ген Е - за нормальное развитие слухового нерва (DdEe). У рецессивных гомозигот dd будет недоразвита улитка, а при генотипе ее - слуховой нерв. Люди с генотипами D-ee, ddE- и ddee будут глухими. У млекопитающих и человека для защиты от вирусов вырабатывается специфический белок интерферон. Его синтез в организме человека обусловлен комплементарным взаимодействием двух неаллельных генов, локализованных в разных (второй и пятой) хромосомах. Гемоглобин человека содержит 4 полипептидные цепи, каждая из которых кодируется отдельным независимым геном. Следовательно, в синтезе гемоглобина участвуют 4 комплементарных гена. II. Один из доминантных комплементарных генов имеет фенотипическое проявление, а второй не имеет; одновременное их присутствие в генотипе обусловливает новый вариант признака. Так у мышей наследуется окраска шерсти агути (в основании и на конце волос — черный пигмент, а в средней части — желтое кольцо). Ген А детерминирует синтез черного пигмента, его аллель а не дает информации для синтеза пигмента. Ген В распределяет пигмент вдоль волоса неравномерно, а его аллель b — равномерно:
Расщепление - в соотношении 9:3:4. III. Каждый из комплементарных генов имеет собственное фенотипическое проявление; одновременное их присутствие в генотипе обусловливает развитие нового варианта признака. Так наследуется форма гребня у кур:
Расщепление - в соотношении 9:3:3:1. При эпистазе доминантный (рецессивный) ген из одной аллельной пары подавляет действие доминантного (рецессивного) гена из другой аллельной пары. Это явление противоположно комплементарности. Подавляющий ген называется супрессором (ингибитором). Различают доминантный и рецессивный эпистаз. Примером доминантного эпистаза может служить полидактилилия. Иногда встречается у «совершенно здоровых» родителей. Предполагается, что действие данного аллеля у родителей подавлялось другими генеми. Примером рецессивного эпистаза является "бомбейский феномен". У женщины, получившей от матери аллель IВ, фенотипиче-ски определялась 1(0) группа крови. При детальном исследовании было установлено, что действие гена IВ (синтез в эритроцитах антигена В) было подавлено редким рецессивным геном, который в гомозиготном состоянии оказал эпистатическое действие. В проявлении некоторых наследственных болезней обмена веществ (ферментопатий) основную роль играет эпистатическое взаимодействие генов, когда наличие или отсутствие продуктов реализации одного гена препятствует образованию активных ферментов, кодируемых другим геном. При полимерии гены из разных аллельных пар влияют на степень проявления одного и того же признака. Полимерные гены принято обозначать одной буквой латинского ал фавита с цифровыми индексами, например A1A1A2а3а3 и т.д. Признаки, детерминируемые полимерными генами, называются полигенными (мультифакториальными). Таким образом у животных и человека наследуются многие количественные и некоторые качественные признаки: рост, масса тела, величина артериального давления, цвет кожи и др. Степень проявления этих признаков зависит от количества доминантных генов в генотипе (чем их больше, тем сильнее выражен признак) и в значительной мере от влияния условий среды. У человека может наблюдаться предрасположенность к различным заболеваниям: гипертонической болезни, ожирению, сахарному диабету, шизофрении и др. Данные признаки при благоприятных условиях среды могут и не проявиться или быть слабовыраженными. Это отличает полигенно-наследуемые признаки от моногенных. Изменяя условия среды и проводя профилактические мероприятия, можно значительно снизить частоту и степень выраженности некоторых мультифакториальных заболеваний. Суммирование "доз" полимерных генов (аддитивное действие) и влияние среды обеспечивают существование непрерывных рядов количественных изменений. Пигментация кожи у человека определяется пятью или шестью полимерными генами. У коренных жителей Африки преобладают доминантные аллели, у представителей европеоидной расы — рецессивные. Мулаты являются гетерозиготами и имеют промежуточную пигментацию. У родителей-мулатов рождаются как белые, так и чернокожие дети. Минимальное количество полимерных генов, при котором проявляется признак, называется пороговым эффектом. Под "эффектом положения" понимают взаимное влияние генов разных аллелей, занимающих близлежащие локусы одной хромосомы. Оно проявляется в изменении их функциональной активности. Резус-принадлежность человека определяется тремя генами, расположенными в коротком плече первой хромосомы на близком расстоянии друг от друга (тесно сцепленными). Каждый из них имеет доминантную и рецессивную аллели (С, D, Е и с, d, e). Организмы с набором генов CDE/cDe и CDe/cDE генетически идентичны (у них общий баланс генов одинаковый). Однако у лиц с первой комбинацией генов образуется много антигена Е и мало антигена С, а у лиц со второй комбинацией аллелей - наоборот, мало антигена Е и много антигена С. Вероятно, близкое соседство аллели Е с аллелью С (первый случай) снижает функциональную активность последней.
Сцепленное наследование В 1908 г. У. Сэттон и Р. Пеннет обнаружили отклонения от свободного комбинирования признаков согласно третьему закону Менделя. В 1911—1912 гг. Т. Морган с соавторами описали явление сцепления генов — совместной передачи группы генов из поколения в поколение. Опыты проводились на мухах дрозофилах с учетом двух пар альтернативных признаков — серый и черный цвет тела, нормальные и короткие крылья. При скрещивании гомозиготных особей с серым телом и нормальными крыльями и особей с черным телом и короткими крыльями получено единообразие первого поколения, особи которого имели доминантные признаки:
Для выяснения генотипа гибридов первого поколения Морган провел анализирующее скрещивание. Он скрестил рецессивную гомозиготную самку с дигетерозиготным самцом (I), затем провел реципрокное скрещивание (II):
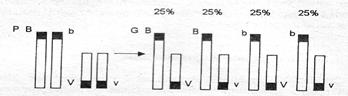
При свободном комбинировании генов, согласно третьему закону Менделя, в поколении и I и II скрещивания должны были появиться мухи четырех разных фенотипов (по 25%). Поясним это схемой, приведенной на рис. 4.3.
Рис. 4.3. Схема свободного комбинирования генов
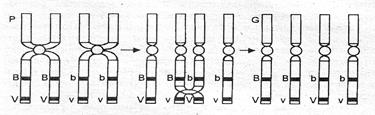
При I скрещивании Морган получил мух только двух фенотипов (по 50%) с признаками родителей. Он пришел к выводу, что гены, детерминирующие цвет тела и длину крыльев, локализованы в одной хромосоме и передаются вместе, т. е. сцепленно. Объяснить это явление можно схемой, приведенной на рис. 4.4.
Рис. 4.4. Схема образования гамет при полном сцеплении
Одна из пары гомологичных хромосом содержит 2 доминантных гена (BV), а другая — 2 рецессивных (bv). В процессе мейоза одна хромосома (с генами BV) попадет в одну гамету, а другая (с генами bv) — в другую. Таким образом, у дигетерозиготного организма образуется не 4, а только 2 типа гамет, и потомки будут иметь такое же сочетание признаков, что и родители. Явление, при котором гены, локализованные в одной хромосоме, всегда передаются вместе, называется полным сцеплением. При изучении результатов II скрещивания было обнаружено нарушение полного сцепления генов. Если дигетерозиготную самку мухи дрозофилы скрестить с рецессивным самцом, то получаются 4 разновидности фенотипов потомков: 41,5% с серым телом и длинными крыльями, 41,5% с черным телом и короткими крыльями и по 8,5% гибридных форм — с серым телом и короткими крыльями и с черным телом и длинными крыльями. В данном случае сцепление оказывается неполным, т. е. происходит перекомбинация генов, локализованных в одной хромосоме. Это объясняется кроссинговером — обменом участками гомологичных хроматид В процессе конъюгации хромосом в профазе мейоза I. Каждая из хроматид попадает в отдельную гамету. Образуется 4 типа гамет, но в отличие от свободного комбинирования их процентное соотношение будет неравным, так как кроссинговер происходит не всегда (рис. 4.5).
Рис. 4.5. Схема образования гамет при кроссинговере
Сила сцепления между генами (частота кроссинговера) зависит от расстояния между ними: чем больше расстояние, тем меньше сила сцепления и тем чаще может происходить кроссинговер. Расстояние между генами определяется в процентах кроссинговера. За единицу его берется 1 % кроссинговера, а сама единица названа морганидой (в честь Моргана). Гаметы, в которые попали хроматиды, не претерпевшие кроссинговера, называются некроссоверными; их обычно больше. Гаметы, в которые попали хроматиды, претерпевшие кроссинговер, называются кроссоверными; их обычно меньше. Итак, если исследуемые гены расположены в разных парах хромосом, происходит их свободное комбинирование согласно третьему закону Менделя. При анализирующем скрещивании мы получим равное количество потомков с различными сочетаниями признаков. Если исследуемые гены локализованы в одной паре гомологичных хромосом и происходит кроссинговер, мы также получим потомков с различными сочетаниями признаков, но количество их будет неравным (рекомбинантных, или кроссоверных, особей будет меньше). Если исследуемые гены локализованы в одной паре гомологичных хромосом и кроссинговер не происходит, то гибридные формы не образуются и потомки будут иметь такое же сочетание признаков, как у родителей. Кроссинговер при образовании гамет происходит у особей обоего пола большинства растений и животных, за исключением самца мухи дрозофилы и самки тутового шелкопряда. Гены, локализованные в одной хромосоме, передаются вместе (сцепленно) и составляют одну группу сцепления. Так как в гомологичных хромосомах локализованы аллельные гены, то группу сцепления составляют две гомологичные хромосомы и количество групп сцепления равно количеству пар хромосом (или гаплоидному их числу). Так, у мухи дрозофилы 8 хромосом — 4 группы сцепления, у человека 46 хромосом — 23 группы сцепления. Перечислим основные положения хромосомной теории наследственности (Т. Морган с соавторами, 1911): 1.Гены расположены в хромосомах линейно в определенных локусах. Аллельные гены занимают одинаковые локусы в гомологичных хромосомах. 2.Гены, расположенные в одной хромосоме, образуют группу сцепления; число групп сцепления равно гаплоидному набору хромосом. 3.Между гомологичными хромосомами возможен обмен аллельными генами (кроссинговер). 4.Процент кроссинговера пропорционален расстоянию между генами; единица расстояния — морганида — равна 1 % кроссинговера. Зная расстояние между генами, можно построить карту хромосомы. Генетическая карта хромосомы представляет собой отрезок прямой, на котором схематически обозначен порядок расположения генов и указано расстояние между ними в морганидах. Она строится на основе результатов анализирующего скрещивания (рис. 4.6).
Рис. 4.6. Схемы генетической (а) к цитологической (б) карт хромосом
Цитологическая карта хромосомы представляет собой фотографию или точный рисунок хромосомы, на котором отмечается последовательность расположения генов. Ее строят на основе сопоставления результатов анализирующего скрещивания и хромосомных перестроек. Например, если хромосома с доминантными генами будет последовательно терять отдельные локусы (при воздействии на нее мутагенов), то в гетерозиготе начнут проявляться рецессивные признаки. Порядок их появления будет указывать на последовательность расположения генов. Картирование хромосом человека связано с определенными трудностями и проводится с использованием методов гибридизации соматических клеток и ДНК. В настоящее время во многих странах продолжает разрабатываться единая международная программа "Геном человека". В начале 2001 г. была полностью расшифрована нуклеотидная последовательность генома человека и выявлена локализация большинства генов. Дальнейшее картирование хромосом человека будет иметь не только важное научное, но и практическое значение: с помощью методов генной инженерии можно будет проводить профилактику и лечение многих наследственных болезней.
ИЗМЕНЧИВОСТЬ Изменчивость — это свойство, противоположное наследственности; оно заключается в способности живых систем приобретать под действием факторов внешней и внутренней среды новые признаки (морфологические, физиологические, биохимические) и особенности индивидуального развития, отличающие их от родительских форм (рис. 5.1).
Рис. 5.1. Классификация типов изменчивости
Генетическая информация определяет степень развития свойств и признаков организма, которые реализуются в определенных условиях среды. Одна и та же наследственная информация в различных условиях проявляется по-разному, Примером могут служить фенотипические различия однояйцевых близнецов, воспитываемых в разных семьях. Окраска шерсти у гималайских кроликов и сиамских кошек зависит от температуры — более темная шерсть растет на участках тела, подверженных охлаждению. Таким образом, наследуется не готовый признак, а определенный тип реакции на воздействия внешней среды. Степень фенотипического проявления данного гена называется экспрессивностью. Она зависит от факторов внешней среды и влияния других генов. Частота проявления гена называется пенетрантностью. Пенетрантность выражается в процентном отношении числа особей, имеющих данный признак, к числу особей, имеющих данный ген. Различная степень пенетрантности и экспрессивности генов имеет большое значение для медицинской генетики. Отягощенная наследственность, наследственная предрасположенность к заболеванию проявляются при воздействии на организм определенных факторов среды. Фенокопия — это явление, при котором признак под действием факторов среды изменяется и копирует признаки другого генотипа. Например, многократный прием алкоголя во время беременности может привести к развитию алкогольной эмбриопатии — комплексу нарушений развития зародыша, копирующему некоторые наследственные синдромы множественных наследственных пороков (синдром Дубовица, болезнь Дауна и др.). Генокопия — это одинаковое фенотипическое проявление мутаций разных генов. Примером генокопий могут служить различные виды гемофилии, которые клинически проявляются снижением свертываемости крови и вызваны недостаточностью восьмого или девятого фактора свертывающей системы (гемофилия А и В соответственно).
Фенотипическая изменчивость Фенотипическая, или модификационная, изменчивость — это изменения фенотипа без изменений генотипа. Она протекает при непосредственном воздействии факторов внешней среды на ферментативные реакции, происходящие в организме, и носит массовый адаптивный (приспособительный) характер. Например, у растения "водяной лютик" листья имеют разнообразную форму: под водой — стреловидные, на границе воды и воздуха — рассеченные, над водой — в виде сплошной пластинки; монозиготные близнецы могут иметь фенотипические различия, если живут в разных условиях. Так как при модификациях не происходит изменения генетического материала, то эта форма изменчивости является ненаследственной. Дарвин называл ее определенной (предсказуемой, или групповой), потому что особи одного вида в одинаковых условиях внешней среды изменяются однотипно. Свойства модификаций: ▪ не наследуются; ▪ предсказуемые; ▪ адаптивные; ▪ носят массовый характер. Границы модификационнной изменчивости называются нормой реакции, которая определяется генотипом. Она может быть узкой, когда признак изменяется незначительно (например, жирность молока), и широкой, когда признак изменяется в широких пределах (например, количество молока).
Генотипическая изменчивость Основные понятия Генотипическая изменчивость — это изменение фенотипа вследствие изменения генотипа. Понятно, что генотипическая изменчивость передается по наследству. Она подразделяется на комбинативную и мутационную.
Комбинативная изменчивость
Комбинативная изменчивость связана с перекомбинацией генов родителей у потомков без изменения структуры генетического материала. Она обусловлена: ▪ перекомбинацией генов при кроссинговере; ▪ независимым расхождением хромосом и хроматид в анафазах мейоза-I и мейоза-П; ▪ случайным сочетанием гамет при оплодотворении! Например, если у родителей I и IV группы крови, то у детей может быть либо II, либо III группа крови:
Разновидностью комбинативной изменчивости является гетерозис — повышение жизнеспособности и увеличение массы различных пород, сортов и даже видов при гибридизации. Явление "гибридной силы" объясняется переходом большинства генов в гетерозиготное состояние, что увеличивает разнообразие белков организма и способствует его лучшей приспосабливаемости к изменяющимся условиям среды.
Мутационная изменчивость Мутации. Мутагенные факторы
Мутация — это скачкообразное, устойчивое изменение генетического материала под влиянием факторов внешней или внутренней среды, передающееся по наследству. Свойства мутаций: ▪ возникают внезапно; ▪ наследуются; ▪ ненаправленны; ▪ могут возникать повторно. Процесс образования мутаций называется мутагенезом, а факторы, их вызывающие, — мутагенами. Мутагены первоначально воздействуют на генетический материал особи, вследствие чего может измениться фенотип. Это могут быть экзомутагены (факторы внешней среды) и эндомутагены (продукты метаболизма самого организма). Мутагенные факторы подразделяют на физические, химические и биологические. К физическим мутагенам относятся различные виды излучений (преимущественно ионизирующие — альфа-, бета-и гамма-лучи, ультрафиолетовые лучи), температура, влажность и др. Механизмы их действия: 1) нарушение структуры генов и хромосом; 2) образование свободных радикалов, вступающих в химическое взаимодействие с ДНК; 3) разрушение нитей ахроматинового веретена деления; 4) образование димеров — соединение между собой ("сшивка") соседних пиримидиновых оснований одной цепи ДНК (чаще Т-Т). К химическим мутагенам относятся: природные органические и неорганические вещества (нитриты, нитраты, алкалоиды, гормоны, ферменты и др.); продукты промышленной переработки природных соединений (угля, нефти); синтетические вещества, ранее не встречавшиеся в природе (пестициды, инсектициды, пищевые консерванты, моющие средства, дезодоранты); лекарственные препараты, которые могут провоцировать врожденные пороки развития (иммуносупрессанты, некоторые антибиотики, наркотические вещества, синтетические кортикостероиды и др.). Химические мутагены обладают большой проникающей способностью, вызывают преимущественно генные мутации и действуют в период репликации ДНК. Механизм их действия: 1) дезаминирование и алкилирование нуклеотидов; 2) замена азотистых оснований их аналогами; 3) ингибиция синтеза предшественников нуклеиновых кислот и др. К биологическим мутагенам относятся: вирусы (краснухи, кори, гриппа); невирусные паразитарные агенты (микоплазмы, бактерии, риккетсии); продукты метаболизма паразитов (токсоплазмы, кошачьего сосальщика, трихинеллы). Невирусные и вирусные_агенты могут быть причиной инфекционного мутагенеза: они способны нарушить течение митоза, вызвать разрывы хромосом и хроматид, встроить свою ДНК в ДНК клеток хозяина. Продукты жизнедеятельности паразитов — возбудителей болезней могут действовать как химические мутагены. Супермутагены — это факторы (чаще химической природы), повышающие частоту мутаций в сотни — десятки тысяч раз (например, колхицин, этиленимин, иприт). Они используются для получения индуцированных мутаций в селекции. Антимутагены значительно снижают частоту мутаций. К ним относятся около 200 природных и синтетических соединений: некоторые аминокислоты (гистидин, метионин и др.); витамины (токоферол, аскорбиновая кислота, каротин и др.); фармакологические средства (интерферон, антиоксиданты, оксипиридины и др.); пищевые продукты (некоторые виды бобов, черный перец, капуста, экстракт яблок). Ряд антимутагенов используют в качестве радиопротекторов.
Классификация мутаций По причинам, вызвавшим мутации, их подразделяют на спонтанные и индуцированные. Спонтанные (самопроизвольные ) мутации происходят под действием естественных мутагенных факторов внешней среды без вмешательства человека, например наследственные болезни обмена веществ. Их причинами являются ошибки репликации и репарации ДНК, действие перекисей и альдегидов, образующихся в организме, различные виды естественных излучений. Индуцированные мутации — результат направленного воздействия определенных мутагенных факторов. Впервые они были получены в 1925 г. Г.А. Надсоном и Г.С. Филипповым при облучении грибов радием. По мутировавшим клеткам мутации подразделяют на гаметические и соматические. Гаметические мутации (генеративные) происходят в половых клетках, проявляются у потомков, передаются по наследству при половом размножении (гемофилия, фенилкетонурия). Соматические мутации происходят в соматических клетках, передаются по наследству только при вегетативном размножении и проявляются у самой особи (разный цвет глаз у одного человека, белая прядь волос, опухоли). По исходу для организма мутации бывают: отрицательные — летальные, несовместимые с жизнью (например, отсутствие головного мозга) и полулетальные — снижающие жизнеспособность организма (например, болезнь Дауна); нейтральные — существенно не влияющие на процессы жизнедеятельности (например, веснушки); положительные — повышающие жизнеспособность (например, появление четырехкамерного сердца в процессе эволюции хордовых животных). Последние возникают редко, но имеют большое значение для прогрессивной эволюции. По изменениям генетического материала мутации подразделяют на геномные, хромосомные и генные.
Геномные мутации
Геномные мутации обусловлены изменениями числа хромосом. К ним относятся полиплоидия, гаплоидия и анеуплоидия. Аномалии числа хромосом могут быть вызваны разными причинами. Наиболее часто геномные мутации являются следствием: нерасхождения хромосом, когда две или несколько гомологичных хромосом остаются соединенными вместе и в анафазу отходят к одному полюсу вследствие разрушения нитей веретена деления; анафазного отставания, когда одна или несколько хромосом в процессе анафазного движения отстают от других. Реже причиной геномных мутаций является полиплоидизация. Полиплоидия — это кратное гаплоидному увеличение числа хромосом в соматических клетках (3n, 4n, 5n,..). Полиплоидия, как правило, используется в селекции растений и приводит к повышению урожайности. У млекопитающих и человека это летальные мутации. Гаплоидия (1n) — одинарный набор хромосом в соматических клетках, например у трутней пчел. Жизнеспособность гаплоидов снижается, так как в этом случае проявляются все рецессивные гены, содержащиеся в единственном числе. Для млекопитающих и человека это летальная мутация. Анеуплоидия — некратное гаплоидному уменьшение или увеличение числа хромосом (2n±1, 2n±2 и т. д.). Существует несколько разновидностей анеуплоидии: ▪ трисомия — три гомологичные хромосомы в кариотипе, например при синдроме Дауна (трисомия по 21-й хромосоме); ▪ моносомия — в наборе одна из пары гомологичных хромосом, например при синдроме Шерешевского — Тернера (моносомия X). Моносомии по первым крупным парам хромосом являются для человека летальными мутациями. Иногда встречаются двойные, тройные (по двум и трем разным хромосомам) моно- и трисомии, тетрасомии и др.; ▪ нулисомия — отсутствие пары хромосом (летальная мутация); у человека неизвестна. Геномные мутации всегда проявляются фенотипйчески и легко обнаруживаются цитогенетическими методами.
Хромосомные мутации
Хромосомные мутации (аберрации) обусловлены изменением структуры хромосом. Они могут быть внутрихромосомными и межхромосомными. К внутрихромосомным мутациям относятся перестройки внутри одной хромосомы. Делеция (нехватка) – отсутствие части хромосомы (частичная моносомия). Выделяют нехватки терминальных и средних участков (рис. 5.2).
Рис. 5.2. Схема внутрихромосомных аберраций
Делеция практически любой части хромосом может нарушить эмбриональное развитие и проявиться множественными врожденными пороками. Например, делеция участка короткого плеча 5-й (5 р -) хромосомы у человека приводит к развитию синдрома "кошачьего крика" (недоразвитие гортани, задержка умственного развития, пороки сердца и др.). При делеции терминальных участков обоих плеч хромосомы (удаляются теломеры) часто наблюдается замыкание оставшейся структуры в кольцо — образование кольцевых хромосом (рис. 5.3).
Рис. 5.3. Схема формирования кольцевых хромосом и фрагментов
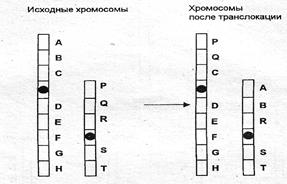
Дупликация (частичная трисомия) – удвоение участка хромосомы. Примером может служить синдром трисомии по короткому плечу 9-й хромосомы (9 р +) у человека, проявляющийся умственной отсталостью, задержкой роста, микроцефалией и другими пороками (см. рис. 5.2). Инверсия – отрыв участка хромосомы, поворот его на 180° и прикрепление к месту отрыва. При этом наблюдается нарушение порядка расположения генов (см. рис. 5.2) Межхромосомные перестройки происходят между негомологичны- ми хромосомами. Транслокация – это обмен сегментами между негомологичными хромосомами. Различают: реципрокные транслокации, когда две хромосомы обмениваются сегментами (рис. 5.4); нереципрокные, когда сегменты одной хромосомы переносятся на другую (рис. 5.5); робертсоновские, когда две акроцентрические хромосомы соединяются своими центромерными районами (рис. 5.6). Иногда может происходить поперечный, а не продольный, как обычно, разрыв хроматид в области центромер; в этом случае образуются изохромосомы (рис. 5.7), представляющие собой зеркальное отображение двух одинаковых плеч (длинных или коротких).
Рис. 5.4. Схема реципрокной транслокации
Рис. 5.5. Схема нереципрокной транслокации
Рис. 5.6. Схема робертсоновской транслокации
Рис. 5.7. Схема образования изохромосом
Нехватки (частичные моносомии) и дупликации (частичные трисомии) всегда проявляются фенотипически, так как изменяется набор генов и нарушается регуляция их активности в процессе эмбриогенеза. Инверсии и транслокации фенотипически проявляются не всегда; они могут быть сбалансированными, когда не происходит ни увеличения, ни уменьшения генетического материала и сохраняется общий баланс генов. При инверсиях и транслокациях, происшедших в клетках половых желез, предшественницах гамет, затрудняется конъюгация гомологичных хромосом, что может служить причиной нарушения распределения генетического материала между дочерними клетками. Хромосомные аберрации выявляются цитогенетическими методами с помощью специальной дифференциальной окраски хромосом.
Генные мутации
Генные (точковые) мутации, или трансгенации, связаны с изменениями структуры гена (молекулы ДНК). Генные мутации могут затрагивать как структурные, так и функциональные гены. Изменения структурных гено |
||||
|
|
Последнее изменение этой страницы: 2016-12-10; просмотров: 526; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.141.198.113 (0.017 с.) |