Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Репродуктивные органы репродуктивной системыСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
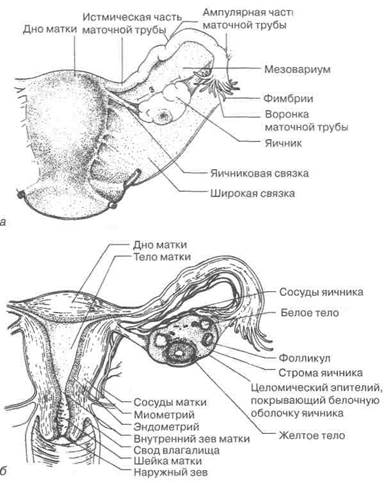
1.3.1. Анатомо-физиологическая и гистофизиологическая характеристика половых органов женщины в репродуктивном периоде 1.3.1.1. Яичники Яичники половозрелой женщины расположены в малом тазу (рис. 1.6), несколько асимметрично на заднем листке широкой связки. Положение яичников в полости малого таза в этом возрасте относительно малоподвижное. Смещение их в брюшную полость наблюдается лишь в период роста организма, а также во время беременности; в конце послеродового периода яичники вновь опускаются в малый таз. Величина и масса тела яичника в возрасте женщины старше 20 лет изменяются незначительно: размер 4,0—4,5 х 2,0—2,5 см (с поперечником 1—2 см), масса 6,0—7,5 г. Правый яичник несколько больше левого. Консистенция гонад плотноватая. Поверхность их до периода половой зрелости гладкая, в репродуктивном периоде становится неровной. Цвет белесоватый, матовый. Яичники не имеют брюшинного покрова; последним покрыта только брыжейка яичника (мезоварий), которая является короткой частью заднего листка широкой связки. К брыжейке яичники прикреплены нижним краем. Каждый яичник имеет две связки: одна из них (воронко-тазовая связка) направляется от верхнего полюса яичника к боковой стенке таза, другая (собственная связка) связывает яичник с маткой, где связка заканчивается позади и несколько ниже маточной трубы. В связках проходят кровеносные и лимфатические сосуды, а также нервы. В яичники основная масса сосудов проходит через мезоварий. Место, где они входят в гонады, именуется воротами яичников. Кровоснабжение гонад осуществляется в основном за счет яичниковой артерии и яичниковой ветви маточной артерии. Кровеносные сосуды в яичниках имеют большое количество анастомозов в корковом и мозговом слоях. Мозговой слой особенно богат сосудами; он граничит с мезоварием. Кровеносные сосуды анастомозируют как в пределах своего слоя, так и между корковым и мозговым слоем, что обеспечивает
Рис. 1.6. Внутренние половые органы женщины в репродуктивном периоде: общий вид (а) и в разрезе (б). Левый яичник, левая маточная труба, матка и влагалище вскрыты; брюшина частично удалена возможность адекватного местного изменения кровообращения [8]. Установлено анастомозирование лимфатической системы яичников, матки и маточных труб с сосудами толстой и прямой кишки, а также аппендикса, мочевого пузыря, почек и надпочечников. Яичники имеют симпатическую адренергическую и парасимпатическую
1 3 Репродуктивные органы репродуктивной системы холинергическую иннервацию. Толстые и тонкие пучки нервных волокон входят через ворота яичников в их мозговой слой как самостоятельно, так и вместе с кровеносными сосудами, образуя вокруг них сплетение. Кровеносные сосуды яичников обильно снабжены нервами и в корковом слое. Соответственно разветвлению сосудов в яичниках происходит деление сопровождающих их нервных стволов. Некоторые из нервных волокон, отделяясь в корковом слое от нервных стволов и соединяясь между собой, образуют на стенках сосудов тончайшие нервные сплетения. Нервные волокна, проходящие в корковом слое, окружают растущие фолликулы (на стадии больших зреющих фолликулов) и зрелый (преовуляторный) фолликул, располагаясь в области внутренней и наружной соединительнотканных оболочек фолликула (текальные оболочки — theca interna и theca externa). Равным образом это относится и к активным в функциональном отношении атретическим фолликулам (атрети-ческие фолликулы с ясно выраженной theca interna). По существующим представлениям, в репродуктивном периоде иннервация зреющих и зрелых фолликулов, атретических фолликулов, активных в функциональном отношении, а также желтых тел является весьма сложной. Яичники покрыты однорядным кубическим эпителием. Наблюдения показывают, что поверхностный эпителий яичников женщин (в отличие от плодовых и детских гонад) в гистологических срезах, как правило, почти отсутствует; чаще он сохраняется в углублениях (бороздках) неровной поверхности яичников. Под поверхностным эпителием располагается белочная оболочка, представленная тонким слоем плотной соединительной ткани, богатой коллагеновыми волокнами. С возрастом она уплотняется. В корковом слое, составляющем большую часть яичников, располагаются многочисленные примордиальные фолликулы (обычно под белочной оболочкой), фолликулы на различных этапах созревания и атрезии, а также желтые тела на разных стадиях развития, в том числе старые желтые тела, относящиеся к предшествующим менструальным циклам. Строма богата округлыми и веретенообразными клетками, которые располагаются в густой сети коллагеновых волокон, и бедна эластическими волокнами. Строма мозгового слоя, в отличие от коркового слоя, бедна клетками, но богата коллагеновыми и эластическими волокнами; в мозговом слое или воротах (в последних — чаще) встречается сеть яичника (rete ovarica), каналь-
Глава 1. Структура и функция репродуктивной системы в возрастном аспекте цы которой выстланы кубическим эпителием, иногда уплощенным. Сеть яичника представляет собой остатки эмбриональных структур мезонефрального происхождения. В области ворот яичника, даже в мозговом слое и в мезоварии, могут встречаться хилюсные клетки (рис. 1.7), являющиеся аналогами клеток Лейдига яичка. Хилюсные клетки находятся в тесном контакте с кровеносными сосудами и нервами. При гистотопогра-фическом исследовании многочисленных ступенчатых срезов они обнаруживаются в воротах яичников в 70—90 % случаев. По наблюдениям L. Нопоге и К. О'Нага [106], в 52 % случаев хилюсные клетки могут быть обнаружены в эндо- и перисальпинксе. Хилюсные клетки (клетки Лейдига) характеризуются полигональной и округлой формой, имеют эозинофильную гранулярную цитоплазму, содержащую бурый пигмент, а также кристаллоиды Рейнке в виде палочковидных образований. При электронно-микроскопическом исследовании установлено, что эти кристаллоиды имеют структуру истинных кристаллов, однако они не выявляются в гистологических срезах значительной части тщательно исследуемых яичников. При обнаружении в цитоплазме хилюсных клеток мелких вакуолей в последних выявляют липиды. Хилюсные клетки могут продуцировать андрогены, но в небольшом количестве. В воротах яичника, хотя и редко, обнаруживаются эмбриональные остатки коры надпочечни-
Рис. 1.7. Хилюсные клетки в воротах яичника, х 350 40 1 3. Репродуктивные органы репродуктивной системы ков (добавочные надпочечники). Они встречаются в любом возрасте в том числе в репродуктивном периоде. Примордиальные фолликулы состоят из яйцеклетки (ооцита) в диктиотене профазы мейоза, окруженной одним рядом уплощенных клеток фолликулярного эпителия (рис. 1.8).
Рис. 1.8, Участок коркового слоя яичника, под белочной оболочкой которого видны примордиальные фолликулы (часть из них без ооцитов) Первичные фолликулы. Увеличение яйцеклетки и округление фолликулярных клеток (последние приобретают кубическую форму) относятся к числу первых признаков начинающегося развития фолликулов. В процессе созревания фолликулов окружающие ооцит клетки увеличиваются в размерах, в них появляются митозы, обусловленные действием ФСГ. Слой фолликулярного эпителия утолщается до 8-10 рядов клеток, превращаясь в малый зреющий фолликул. Вторичные фолликулы. На этом этапе созревания фолликула происходит дальнейшее нарастание количества рядов фолликулярного эпителия (гранулезная оболочка), в толще которого начинается формирование полости. В этот период, с началом превращения малого зреющего фолликула в большой зреющий фолликул (рис. 1.9), образовавшиеся текальные оболочки (theca interna и theca externa) отчетливо дифференцируются, а яйцеклетка окружена прозрачной зоной (zona pellucida), представленной гликозаминогликанами. В отличие от первичного фолликула, формирование и последующая дифференциация вторичного фолликула полностью зависимы
Рис. 1.9. Участок стенки вторичного фолликула на этапе превращения его в большой зреющий фолликул; theca interna лютеинизирована, х 300 от ЛГ и ФСГ [44]. По мере дальнейшего созревания фолликула и увеличения количества жидкости в фолликулярной полости (главным образом, за счет секреции клеток гранулезы) ооцит, непосредственно окруженный гранулезой, расположенной радиально в виде лучистого венца, оказывается сдвинутым к периферии фолликула, образуя таким образом яйценосный бугорок — cumulus oophorus. Среди гранулезы вторичного фолликула иногда обнаруживаются микрополости — тельца Калла—Экснера. Theca interna вторичных и, особенно преовуляторных фолликулов, богато васкуляризована, содержит липиды. Преовуляторный фолликул представляет собой зрелый фолликул, полость которого в этот период является наибольшей, а сам он смещен ближе к поверхности яичника. В яйцеклетке заканчивается 1-е деление созревания, отделяется 1-е полярное тельце, и ооцит переходит во 2-е деление созревания, которое «застывает» на мета-фазе до оплодотворения [8]. Гранулезная оболочка преовуляторного фолликула (вне яйценосного бугорка) у человека состоит всего из 2—4 рядов клеток фолликулярного эпителия в отличие от зрелого фолликула яичника животных [31]. Яйцеклетка в преовуляторном фолликуле окружена 3—4 слоями (рядами) плотно расположенных клеток лучистого венца. Ко времени овуляции клетки в этой области разрыхляются, уве- 1.3. Репродуктивные органы репродуктивной системы личиваются и межклеточные пространства. Часть клеток отрывается и лизируется. На вершине преовуляторного фолликула (расположенного на уровне поверхности органа) формируется небольшая бессосудистая область — стигма. Перед овуляцией стенка фолликула в области стигмы представлена одним рядом клеток гранулезы, прилежащих непосредственно к поверхностному эпителию. Приведенные данные основаны на результатах исследования яичников животных (мыши), полученных путем применения сканирующей электронной микроскопии [2]. До настоящего времени существуют различные гипотезы относительно возможных механизмов овуляции. Длительное время существовало убеждение, что в момент овуляции стенка зрелого фолликула в результате увеличения размеров последнего и нарастания внутрифолликулярного давления разрывается и яйцеклетка вместе с фолликулярной жидкостью попадает на фимбрии, а затем в ампулярную часть маточной трубы. В последние годы высказаны сомнения относительно этой чисто механической теории и выдвинуты иные представления, часть которых обобщена Б. И. Желез-новым [36], О. В. Волковой и Н. С. Миловидовой [8]. К их числу можно отнести предположение о ведущей роли протеолитических ферментов в процессе разрыва фолликулов, гипотезы о возможном значении иммунных реакций в механизмах развития овуляции [88] и о нервно-мышечном механизме последней.
Согласно одной из последних гипотез, зрелый фолликул человека и млекопитающих содержит гладкие мышечные клетки или клетки, которым свойственны многие характерные особенности гладких миоцитов. Это доказано методами электронной микроскопии (обнаружение миофиламентов) и иммуногистологическими исследованиями (выявление сократительных белков — актина и миозина). Эти клетки, расположенные в волокнистой соединительной ткани theca externa, не имеют отношения к сосудам. По мнению некоторых авторов, сократительная деятельность стенки фолликула обеспечивается как адренергическими а-рецепторами, так и холинергическими рецепторами, а расслабление стенки фолликула осуществляется через в2-адренорецепторы. Эти механизмы контрактильной деятельности способны оказывать влияние на внутрифолликулярное давление и процесс овуляции непосредственно. После овуляции полость фолликула спадается, а стенки его приобретают вид фестонов; на этом месте в последующем развивается желтое тело. Глава 1. Структура и функция репродуктивной системы в возрастном аспекте Принято считать, что спонтанной циклической овуляции подвергается лишь один зрелый фолликул. Вместе с тем наблюдения показывают, что в яичнике иногда можно обнаружить более чем одно свежее желтое тело. Этот факт объясняют следствием овуляции более одного фолликула в различные сроки одного и того же цикла. Причины неодновременного формирования двух доминантных фолликулов в течение одного менструального цикла остаются не до конца ясными и в настоящее время, однако сам факт является доказанным [147]. Об этом свидетельствуют и указания, согласно которым овуляция бывает дважды в одном и том же месяце (по одному разу в каждом яичнике) примерно 1 раз в 7,5 лет. Таким образом, в течение менструального цикла в яичниках женщины развивается несколько фолликулов, хотя лишь один (редко — два) достигает стадии зрелого фолликула. Остальные начавшие развитие фолликулы чаще подвергаются кистозной, а затем и облитерационной атрезии с исходом в фиброзные (гиалиновые) атретические тела. В theca interna фолликулов, находящихся на стадии атрезии, как и в больших зреющих и зрелых фолликулах, содержатся липиды. Theca interna атретических фолликулов может быть с признаками гиперплазии и/или лютеинизации. При значительных скоплениях лютеинизированных клеток, имеющих отношение к theca interna, они именуются интерстициальной железой. По современным представлениям, интерстициальная железа является эндокринным образованием и, подобно theca interna фолликулов, продуцирует эстрогены. Высказываются предположения о том, что в репродуктивном периоде интерстициальная железа служит важным источником секреции эстрогенов в постовуляторной фазе каждого менструального цикла [45].
Как известно, в развитии свежего желтого тела различают три стадии: пролиферацию, васкуляризацию и расцвет. На стадии пролиферации происходят размножение гранулезы и лютеинизация клеток theca interna. Спустя несколько часов начинается лютеинизация гранулезы. Однако к этому времени она не достигает той выраженности, которая свойственна текалютеиновым клеткам. На стадии васкуляризации в гранулезолютеиновую ткань врастают тонкостенные кровеносные сосуды theca interna. Последние продвигаются в направлении сохраняющейся центральной полости желтого тела, обычно содержащей сгусток крови (так называемое центральное ядро желтого тела). Капилляры оплетают каждую клетку желтого I 3. Репродуктивные органы репродуктивной системы тела и вместе с сопровождающей их соединительной тканью окружают центральное ядро. Клетки гранулезы становятся крупными, полигональными. Цитоплазма их ацидофильная. В мелких темных ядрах лютеиновых клеток митозы обнаруживаются до 18-го цикла. На стадии расцвета желтого тела оно является окончательно сформированным и достигает величины 1,0—1,5 см. Клетки его резко увеличены в размере, богаты цитоплазмой, содержащей мельчайшие капельки липидов и липохромный пигмент, ядра крупные везикулярные с наличием ядрышек. Гранулезолютеиновые клетки, в отличие от текалютеиновых, продуцируют прогестерон. Текалютеиновые клетки, располагающиеся по периферии желтого тела (по величине они меньше лютеиновых), продуцируют эстрогены. В стадии расцвета желтое тело человека находится 10—12 дней. Дистрофические изменения в лютеиновых клетках — начало обратного развития желтого тела. Они проявляются в виде вакуолизации, накопления крупнокапельного нейтрального жира, пикноза ядер, уменьшения размера клеток, врастания соединительной ткани с замещением распадающихся лютеиновых клеток. По данным, приводимым в литературе, субмикроскопически в лютеоцитах регрессирующего желтого тела отмечаются дезорганизация цитоплазма-тической сети и матрикса, образование аутофагических вакуолей и миелиновых структур, повышение количества липидов и лизосом. Принято считать, что обратное развитие желтого тела начинается в конце менструального цикла, однако, по данным Б. И. Желез-нова [36], Е. Novak и J. Woodruff [142], A. Blaustein [144], регрессия желтого тела может начинаться и с 21—23-го дня цикла. Обратное развитие желтого тела продолжается не менее 2 мес, заканчиваясь формированием белого тела, которое представляет собой гиалиновое образование. Яичники имеют хорошо развитую не только кровеносную, но и лимфатическую систему. Особенности и степень васкуляризации фолликулов зависят от развития theca interna как в растущих, так и активных в функциональном отношении атретических фолликулах. При дифференцировке в зреющем фолликуле текальных оболочек theca interna (по сравнению с theca externa) особенно богата капиллярами; так же распределяются и лимфатические микрососуды. Согласно приводимым в литературе данным, наиболее выраженная васкуляризация отмечена у преовуляторных фолликулов, что может быть обусловлено увеличением не только новообразованных, но и Глава 1. Структура и функция репродуктивной системы в возрастном аспекте функционирующих капилляров. Овуляторный процесс начинается с расширения и переполнения кровеносных сосудов микроокружения фолликула. На первых этапах развития желтого тела отмечается обилие широких переполненных кровью капилляров на периферии theca interna. В начальной стадии атрезии фолликулов появляется характерная реакция микрососудистого русла в виде расширения капилляров и переполнения их кровью. В процессе дальнейшей атрезии фолликулов наряду с капиллярами, не имеющими структурных нарушений, выявляются капилляры с явными признаками деструкции. При атрезии фолликулов, имеющих выраженный слой theca interna, их клетки разрастаются параллельно с капиллярами внутренней текальной оболочки [8]. В целом циклически повторяющиеся процессы развития фолликулов, заканчивающиеся овуляцией с последующим образованием желтого тела, сопровождаются новообразованием мелких кровеносных сосудов и микрососудов — капилляров. После регрессии желтого тела капилляры постепенно запустевают. 1.3.1.2. Маточные трубы Маточные трубы половозрелой женщины, расположенные в малом тазу (почти поперек последнего), покрыты брюшиной. По нижнему краю каждой из труб складка брюшины, представляющая собой верхнюю часть широкой связки, образует брыжейку — мезосаль-пинкс. В маточных трубах самой узкой и короткой является интраму-ральная или интерстициальная часть, которая проходит через стенку матки. К ней примыкает истмическая часть (перешеек), имеющая длину примерно 2-3 см. За перешейком следует ампулярная часть (она составляет около 2/3 всей длины трубы), переходящая в воронку, края которой снабжены бахромками (фимбриями); одна из них (fimbria ovarica) проходит по краю брыжейки, достигая яичника. Длина маточных труб у женщин репродуктивного возраста варьирует от 9 до 13 см, составляя в среднем 10—11 см. В поперечном сечении стенка трубы состоит из трех оболочек: серозной, мышечной и слизистой. Серозная оболочка представлена мезотелием, под которым располагаются в небольшом количестве соединительная ткань, мышечные волокна и кровеносные сосуды. Артериальное кровоснабжение имеет двоякое происхождение. Оно обеспечивается за счет трубных и яичниковых ветвей маточной артерии, про- I 3. Репродуктивные органы репродуктивной системы ходящих в мезосалышнксе; имеются сведения, что трубная ветвь маточной артерии анастомозирует с трубной ветвью яичниковой артерии. Анастомозирующие ветви маточной и яичниковой вен проходят параллельно артериальным сосудам, локализуясь также в мезосалышнксе. Лимфатические сосуды сопровождают кровеносные, главным образом овариальные, сосуды. Мышечная оболочка трубы состоит из гладких мышц и обычно представлена двумя слоями: наружным продольным и внутренним циркулярным; однако в интрамуральной части мышечная оболочка, достигающая почти 1 см толщины, имеет и внутренний продольный слой. Наиболее тонкой мышечная оболочка бывает в ампулярной части (около 0,1 см), в истмусе она достигает толщины около 0,5 см. Количество сосудов в мышечной оболочке возрастает по мере приближения к ампуле. Особенно ими богата воронка. В репродуктивном периоде маточные трубы обладают выраженными сосудистыми приспособлениями для депонирования шунтирования кровотока. По данным Б. И. Глуховца и соавт. [12], депонирование крови обеспечивается множественными посткапиллярными синусами и запирательными венулами, характерными для фимбри-ально-ампулярного отдела труб. Основной механизм шунтирования кровотока в маточных трубах представлен замыкательными артериями и артериоло-венулярными анастомозами, расположенными в наружном слое стенки ампулярно-истмической части. Кроме того, на протяжении всей трубы определяются артериоло-венулярные полушунты, или «ложные анастомозы», являющиеся дополнительным путем для юкстакапиллярного кровотока. Маточные трубы иннервируются от ветвей тазового и яичникового сплетений. Основное нервное сплетение в трубах образуется в субсерозном их слое, откуда нервные волокна идут в направлении серозной и мышечной оболочек органа. Каждая из маточных труб имеет симпатическую и парасимпатическую иннервацию. Слизистая оболочка (эндосальпинкс) образует широкие продольные складки, представлена однорядным цилиндрическим эпителием и, в небольшом количестве, рыхлой волокнистой соединительной тканью (собственной пластинкой), содержащей сосуды, соединительнотканные клетки веретенообразной формы и нервные окончания; в небольшом количестве в ней встречаются гистиоциты, лимфоциты и лаброциты, а также единичные лейкоциты и плазматические клетки. Клеточные элементы преобладают Глава 1. Структура и функция репродуктивной системы в возрастном аспекте в строме слизистой оболочки перешейка, и наоборот, волокнистых структур в ампулярной части больше, чем в истмусе. В каждом из отделов маточных труб эндосальпинкс имеет характерное строение: в ампулярной части складки слизистой оболочки высокие, характеризуются выраженной ветвистостью, в перешейке они ниже и лишены вторичного ветвления, в интрамуральной части складки эндосальпинкса небольшие, в количестве 5—6, а иногда и вовсе отсутствуют. По современным представлениям, особенности строения складок слизистой оболочки в каждом из отделов маточных труб связаны с их функцией. Так, по-видимому, фимбриям свойственна функция захватывания яйцеклетки, а сложный ветвистый рельеф складок эндосальпинкса в ампулярной части препятствует нидации ооцита и в то же время способствует оплодотворению, которое совершается именно в ампуле трубы; что касается складок слизистой оболочки перешейка, то ее функциональная роль проявляется в секреции веществ, необходимых для жизнедеятельности яйцеклетки. Имеются данные, что область перешейка является зоной активной секреции [18, 21]. Прежде чем перейти к освещению структурных и морфофунк-циональных особенностей трубного эпителия, следует остановиться на некоторых изменениях в состоянии волокнистых структур и сосудов, на клеточных реакциях и других их проявлениях. Не углубляясь в анализ структурной перестройки и морфофункциональных изменений стромы и сосудов, приведем лишь некоторые факты, свидетельствующие о значимости их не только в научном, но и практическом отношении. Так, по данным, приводимым О. В. Волковой [8], функциональное состояние системы микроциркуляции подчинено гормональным влияниям. К моменту овуляции постепенно нарастает извитость артериол, наблюдаются расширение капилляров, отек стромы и резкое расширение лимфатических микрососудов. В лютеиновой фазе отмечается, по выражению автора, «нормализация» показателей микроциркуляции. Как показывают наши наблюдения, в конце менструального цикла вновь возникает гиперемия и резко расширяются лимфатические сосуды (рис. 1.10), развивается отек стенки органа. В дифференциальной гистологической диагностике играют роль изменения, обнаруживаемые в маточных трубах во время менструации или вскоре после нее: разволокнение стромы эндосальпинкса при наличии в последнем расширенных лимфатических и кровеносных сосудов, немногочисленных лимфоцитов 1.3. Репродуктивные органы репродуктивной системы и единичных плазматических клеток и лейкоцитов [14]; очаговые скопления последних в просвете маточных труб или одной из них в подобных наблюдениях не такое уж редкое явление (рис. 1.11). Установлен факт циклических колебаний лаброцитов в стенке маточных труб на протяжении менструального цикла. Содержание гликозаминогликанов и кислых гликозаминоглю-куронгликанов в строме эндосальпинкса невелико. Несмотря на
Рис. 1.10. Участок маточной трубы в истмико-ампулярной части ее, местами с резким расширением лимфатических сосудов в складках слизистой оболочки, х 65
Рис. 1.11. Фрагмент эндосальпинкса ампулярной части с различными типами трубного эпителия; преобладают мерцательные клетки в фолликулиновую фазу цикла, х 400 Глава 1. Структура и функция репродуктивной системы в возрастном аспекте относительно небольшое количество стромальных клеток (количество их меньше, чем в строме эндометрия), очаговая децидуальная трансформация слизистой оболочки маточных труб наблюдается во время беременности и в раннем послеродовом периоде примерно в 8 % наблюдений. Таким образом, стромальный и сосудистый компоненты подвергаются различным гистофизиологическим изменениям. Равным образом это относится и к трубному эпителию, состояние которого характеризуется значительным многообразием, отражающим функциональное состояние основных клеточных его типов. Последним свойственны различные структурно-функциональные изменения: они наблюдаются, например, в клетках эпителия маточных труб в различные фазы менструального цикла, во время беременности и лактации, а также в постменопаузе. В трубном эпителии у женщин репродуктивного возраста можно различить четыре основных типа клеток: 1) мерцательные (реснитчатые); 2) секреторные; 3) базаль-ные {интраэпителиалъные пузырчатые, индифферентные); 4) вставочные, или штифтообразные [11, 42, 44, и др.]. Вместе с тем до настоящего времени сохраняются разногласия в оценке клеточного состава трубного эпителия как относительно числа типов клеток, так и их функции. А. Хэм и Д. Кормак [45] выделяют лишь два основных типа клеток трубного эпителия: реснитчатые и секреторные. Именно эти два типа эпителиальных клеток выявлены при применении сканирующей электронной микроскопии О. В. Волковой и соавт. [7]. Обсуждается вопрос и о степени самостоятельности мерцательного и секреторного типов клеток и возможности их взаимного превращения [8]. Мерцательные клетки, как и секреторные, обнаруживаются на всем протяжении маточных труб (см. рис. 1.11). Распределение этих видов клеток в различных частях трубы обратно пропорционально друг другу: количество мерцательных клеток, наиболее многочисленных в фимбриях (85 %), по направлению к маточному концу трубы постепенно снижается, а количество секреторных клеток, наоборот, увеличивается. Мерцательные клетки отличаются от секреторных не только наличием ресничек (около 50%), но и другими цитологическими особенностями: они шире, чем секреторные, а округлое ядро их располагается в центре клетки. Образованию и росту ресничек мерцательных клеток, как и увеличению высоты последних, способствуют эстрогены. Прогестерон стимулиру- 1 3 Репродуктивные органы репродуктивной системы ет секрецию секреторных клеток. Секреторные клетки (они, как и мерцательные, имеют цилиндрическую форму) характеризуются более выраженной базофилией цитоплазмы, содержащей большее количество органелл, чем в мерцательных клетках; их овоидные ядра вытянуты вдоль клетки. Базалъные клетки, располагающиеся на базальной мембране, отличаются округлой формой, светлой цитоплазмой и круглым темноокрашенным ядром. Штифтообразные клетки имеют палочковидное ядро, скудную цитоплазму. В 1-й половине менструального цикла поверхность эпителиального покрова маточных труб ровная. Высота секреторных и мерцательных клеток в начале фолликулиновой фазы обычно не превышает 20 мкм, к моменту овуляции достигает 30—35 мкм. Базальные клетки в фолликулиновой фазе лишь единичные, высотой до 8 мкм, а штифтообразные в это время отсутствуют. Существует мнение, что физиологическая регенерация мерцательных и секреторных клеток осуществляется за счет амитотического деления базальных клеток. Однако нельзя игнорировать и митотическое деление. В наших наблюдениях митозы, хотя и редко, обнаруживались в базальных клетках не только при гиперпластических процессах трубного эпителия, но и в неизмененных или малоизмененных трубах у женщин репродуктивного возраста с двухфазным менструальным циклом. Наличие митозов в базальных клетках трубного эпителия, главным образом в поздней стадии пролиферации, отмечают в последние годы Б. И. Глуховец и соавт. [12]. На протяжении 2-й половины менструального цикла высота эпителиальных клеток, главным образом мерцательных, снижается до 18—20 мкм. Секреторные клетки приобретают разнообразную форму, преимущественно бокаловидную или грушевидную, узким концом обращенную в сторону базальной мембраны. Апикальная часть секреторных клеток выступает над поверхностью мерцательных клеток, вследствие чего поверхность эпителия слизистой оболочки труб становится неровной. Среди секреторных и мерцательных клеток эпителия можно обнаружить штифтообразные клетки. К концу лютеиновой фазы цикла количество базальных и штифтообразных клеток возрастает. Принято считать, что штифтообразные клетки представляют собой дистрофически измененные мерцательные и секреторные клетки. Мерцательные клетки в процессе превращения в штифтообразные уменьшаются в поперечном размере, ядра их становятся пикнотичными, палочковидной формы, реснички исчезают. Образование штифтообразных Глава 1. Структура и функция репродуктивной системы в возрастном аспекте клеток из секреторных происходит после выделения последними секреторного материала. В дальнейшем штифтообразные клетки подвергаются аутолизу, хотя отдельные дистрофически измененные клетки обнаруживаются и в просвете маточных труб. О циклических процессах в маточных трубах свидетельствуют и морфофункциональные изменения трубного эпителия в различные фазы менструального цикла. По данным Н. И. Кондрикова [21], на протяжении менструального цикла в трубном эпителии отмечаются циклические изменения содержания и распределения гликогена, РНК, белков, липидов и активности щелочной и кислой фосфатаз. Уменьшение содержания гликогена в апикальной части мерцательных клеток после овуляции свидетельствует о связи этих изменений с нарастающей мерцательной акгивностью трубного эпителия. Известно, что функция секреторных клеток сводится к выработке веществ, необходимых для жизнеспособности яйцеклетки. В просвете трубы содержится некоторое количество жидкости — секрета, включающего гликопротеины, кислые гликозаминоглюкуронглика-ны, простагландин F2oc, электролиты и различные ферменты, обеспечивающие оптимальную среду для сперматозоидов и яйцеклетки. Максимальная активность секреторных клеток приходится на лютеиновую фазу. Мерцательным клеткам отводится важное место в обеспечении продвижения яйцеклетки по трубе в матку. Однако не менее важным фактором является также перистальтика самой маточной трубы вследствие активных сокращений ее гладких мышц. Принято считать, что перистальтика маточной трубы наиболее выражена в период овуляции. Поскольку одной из основных функций этого органа является захват яйцеклетки после овуляции, воронка трубы соприкасается в момент овуляции с яичником, что обеспечивается сокращениями мускулатуры трубы. Определенная роль в транспорте овулировавшей яйцеклетки к фимбриальному концу маточной трубы отводится сократительной активности трубно-яичниковой связки. В период овуляции спонтанная сократительная ее активность возрастает в 3 раза в сравнении с поздней фолликулиновой и предменструальной фазами, в 6 раз — с ранней фолликулиновой и в 2 раза — с лютеиновой фазой цикла. Под влиянием простагландина F2a и ацетилхолина сократительная активность трубно-яичниковых связок возрастает в 3 раза. Таким образом, у женщин с нормальным менструальным циклом циклические структурные и морфофункциональные измене- 1 3 Репродуктивные органы репродуктивной системы ния трубного эпителия способствуют созданию условий для оплодотворения и начальных этапов развития оплодотворенной яйцеклетки. 1.3.1.3. Матка Матка представляет собой мышечный орган грушевидной формы, расположена в середине полости малого таза в положении antev-ersio-flexio. В матке различают дно, тело, перешеек (истмическую часть) и шейку матки. Дном матки называют верхнюю часть ее, расположенную в области отхождения маточных труб. Матка рожавшей, женщины больше, чем нерожавшей; и имеет массу от 50 до 90 г. Общая длина матки 7—9 см, из которой длина шейки матки составляет 2,5-3,0 см. Соотношение длины тела и шейки матки составляет в среднем 2:1. Полость тела матки имеет треугольную форму, переднезадний диаметр которой 2,5-3,5 см, а наибольшая ширина варьирует от 4,5 до 6 см. В положении anteversio-flexio матка удерживается благодаря связочному аппарату. Круглые связки (в их состав входит гладкая мышечная ткань) отходят от углов матки, располагаясь ниже и несколько кпереди от маточных труб: они проходят в паховые каналы через внутренние и наружные отверстия их и оканчиваются в клетчатке лобка и больших половых губ. Широкие связки матки представляют собой дупликатуру брюшины, идущую от боковых поверхностей матки до боковых стенок таза. Для фиксации матки в типичном нормальном положении круглые и широкие связки не имеют столь существенного значения, как кардинальные связки, представляющие собой мощные пучки соединительной ткани и небольшие пучки гладкомышечной ткани; те и другие расположены в основании широких связок. Крестцово-маточные связки отходят от задней поверхности шейки в области перешейка и идут к прямой кишке (частично вплетаясь в мышечную стенку последней) и крестцу. Опорным аппаратом является тазовое дно. При нормальном положении матка подвижна, легко смещается при изменении состояния соседних органов — при переполнении мочевого пузыря и прямой кишки. На положение матки оказывают влияние положение тела женщины, наличие беременности, подъем тяжестей и ряд других факторов. Кровоснабжение матки обеспечивается маточными и яичниковыми артериями, которые соединены между собой многочис- Глава 1. Структура и функция репродуктивной системы в возрастном аспекте ленными анастомозами. Источником кровоснабжения дна матки являются также артерии круглых связок, в каждой из которых кроме артерии, проходят вена и лимфатические сосуды. В основном кровь к матке, по-видимому, поступает по маточным артериям. Главные стволы маточной артерии располагаются по боковым поверхностям матки, извиваясь и образуя многочисленные ветви, идущие к передней и задней стенкам матки. Последние, анастомо-зируя между собой и ветвями маточной артерии противоположной стороны, образуют богатое сосудистое сплетение в области дна и тела матки. В области угла, образованного трубой и краем матки, восходящая ветвь маточной артерии делится на две (иногда — на три) ветви, из которых одна идет к дну матки, другая — в брыжейку яичника. Восходящая ветвь маточной артерии у рожавших женщин штопорообразно извита, у нерожавших имеет вид дугообразной линии. Факту штоп
|
||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-26; просмотров: 1206; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.133.149.29 (0.021 с.) |