Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Медиаторы ноцицептивной системыСодержание книги
Поиск на нашем сайте С помощью медиаторов ноцицептивной системы информация передается с клетки на клетку. § Субстанция Р (от англ. pain – «боль») – главная. § Нейротензин. § Брадикинин. § Холецистокинин. § Глютамат.
22. – Теории возникновения боли. Механизм возникновения боли согласно теории воротного контроля. Механизмы функционирования антиноцептивной системы.
Теории возникновения боли. Теория специфичности утверждает, что боль представляет собой отдельную сенсорную систему, в которой любой повреждающий стимул активирует специальные болевые рецепторы (ноцицепторы), передающие болевой импульс по специальным нервным путям в спинной мозг и в болевые центры головного мозга, вызывая ответную защитную реакцию, направленную на удаление от раздражителя.
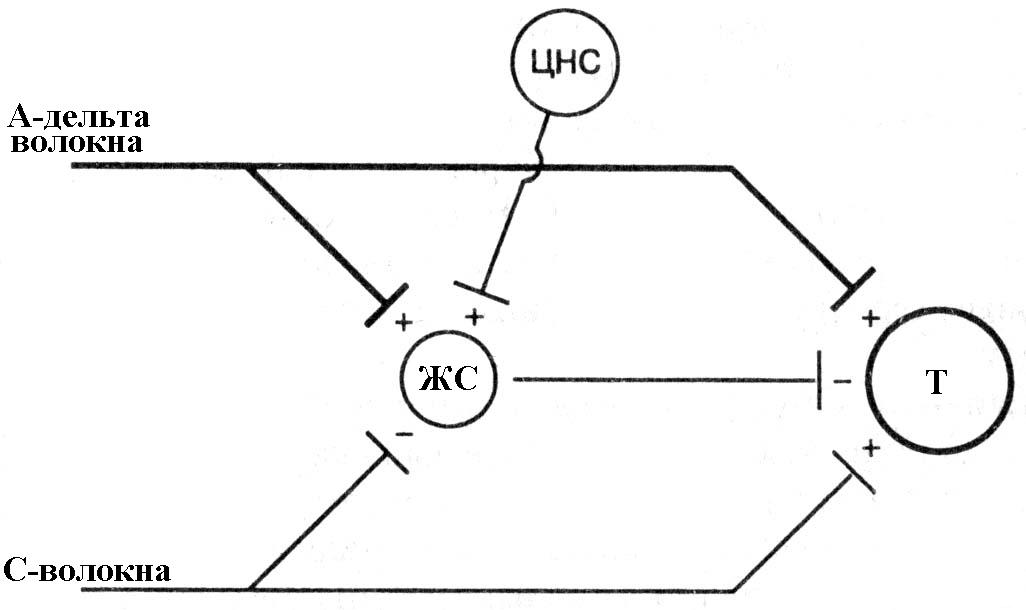
Основой для создания в теории специфичности послужило учение французского философа и физиолога Р. Декарта о рефлексе. В 20-ом столетии правомерность концепции боли, как специфической проекционной сенсорной системы, была подтверждена многочисленными исследованиями и открытиями в анатомии и экспериментальной физиологии. Были обнаружены болепроводящие нервные волокна и болепроводящие пути в спинном мозге, болевые центры в различных отделах головного мозга, медиаторы боли (брадикинин, субстанция Р, ВИП и др.). Согласно теории специфичности, психологическое ощущение боли, её восприятие и переживание признаются адекватными и пропорциональными физической травме и периферическому повреждению. В практической медицинской деятельности это положение привело к тому, что пациенты, страдающие болью и не имеющие очевидных признаков органической патологии, стали считаться "ипохондриками", "невротиками" и, в лучшем случае, направлялись на лечение к психиатру или психотерапевту. Теория интенсивности утверждает, что ощущение боли возникает при раздражении любого рецептора избыточным стимулом (шум, свет). Теория воротного контроля (Melzack, Wall, 1965). Поток болевой импульсации с периферии идет в задний рог спинного мозга по большим миелинизированным (А-дельта) и малым немиелинизированным (С-волокнам) нервным волокнам. Оба типа волокон образуют синапсы с нейронами второго порядка (Т) ("передача/проекция"). Когда Т-нейроны активированы, они поставляют ноцицептивную информацию в мозг. Периферические нервные волокна также образуют синапсы с интернейронами желатинозной сустанции (ЖС), которые при стимуляции угнетают Т-нейроны. А-дельта волокна стимулируют, а С-волокна угнетают интернейроны ЖС, соответственно снижая и повышая центральную передачу ноцицептивных входящих сигналов. Кроме того, стимуляция интернейронов ЖС на подавление активности Т-нейронов происходит через нисходящие пути, начинающиеся в центральной нервной системе (это происходит при активации различными факторами). Баланс между возбуждающими и угнетающими сигналами определяет степень передачи ноцицептивной информации в головной мозг («+» - возбуждающий сигнал; «-» - угнетающий сигнал).
Рис. 8.2. Схема теории «воротного контроля» по R. Melzack, 1999 (объяснение в тексте). Примечание. ЖС - желатинозная субстанция задних рогов спинного мозга, Т - трансмиссивные нейроны.
Основное научно-медицинское значение теории "входных ворот" заключалось в признании спинного и головного мозга активной системой, фильтрующей, отбирающей и воздействующей на входные сенсорные сигналы. Теория утвердила центральную нервную систему ведущим звеном в болевых процессах.
Теория «генератора патологически усиленного возбуждения» в центральной нервной системе подчеркивает значение центральных механизмов в патогенезе боли и определяет роль периферических факторов. Генератор патологически усиленного возбуждения (ГПУВ, генератор) - это агрегат гиперактивных нейронов, продуцирующий чрезмерный неконтролируемый поток импульсов. ГПУВ образуется в поврежденной нервной системе из первично и вторично измененных нейронов и представляет собой новую, необычную для деятельности нормальной нервной системы патологическую интеграцию, возникающую на уровне межнейрональных отношений. Особенностью генератора является его способность развивать самоподдерживающуюся активность. ГПУВ может образовываться практически во всех отделах ЦНС, его формирование и деятельность относятся к типовым патологическим процессам. При создании генератора в системе болевой чувствительности появляются различные болевые синдромы: болевой синдром спинального происхождения (генератор в дорсальных рогах спинного мозга), тригеминальная невралгия (генератор в каудальном ядре тройничного нерва), таламический болевой синдром (генератор в ядрах таламуса). Невромы, повреждения нервов, смещения межпозвонковых дисков вызывают боль и приводят к возникновению патологических центральных процессов. В ЦНС формируется "генератор патологически усиленного возбуждения", в результате значение периферических факторов уменьшается. Поэтому при сильной фантомной невралгической и поясничной боли после удаления невром нервов, грыж диска и т.п. устранение периферических факторов может не привести к прекращению боли.
Возникновение генератора начинается либо с первичной гиперактивации нейронов, либо с первичного нарушения их торможения. При первичной гиперактивации нейронов тормозные механизмы сохранены, но они функционально недостаточны. В этом случае имеет место вторичная недостаточность торможения, которая возрастает по мере развития генератора, при преобладании возбуждения. При первичной недостаточности тормозных механизмов появляются растормаживание и вторичная гиперактивация нейронов. Первичная гиперактивация нейронов возникает вследствие усиленных и длительных возбуждающих воздействий: при синаптической стимуляции, при действии возбуждающих аминокислот, К+ и др. Роль синаптической стимуляции отчетливо видна на примере формирования генератора в ноцицептивной системе. Хронически раздражаемые рецепторы в тканях, эктопические очаги в поврежденных нервах, неврома (хаотически разросшиеся афферентные волокна) являются источником постоянной импульсации. Под влиянием этой импульсации в центральном аппарате ноцицептивной системы формируется генератор. Первичное нарушение торможения нейронов формируется в условиях действия веществ, избирательно повреждающих тормозные процессы. Такой эффект имеет место при действии столбнячного токсина, нарушающего выделение пресинаптическими окончаниями тормозных медиаторов; при действии стрихнина, блокирующего глициновые рецепторы на постсинаптических нейронах спинного мозга, где глицин оказывает тормозной эффект; при действии некоторых конвульсантов, нарушающих постсинаптическое торможение.
Поскольку деятельность генераторных механизмов определяется множественными взаимодействиями, влиять на нее можно путем одновременного использования антидепрессантов, раздражения триггерных точек электрическим током, физиотерапией и др.
Понятие антиноцицептивной системы. Ее уровни, медиаторы.
Антиноцицептивная система Комплекс ноцицептивной системы в равной степени сбалансирован в организме комплексом антиноцицептивной системы, обеспечивающей контроль за активностью структур, участвующих в восприятии, проведении и анализе болевых сигналов. В настоящее время установлено, что болевые сигналы, поступающие с периферии, стимулируют активность различных отделов центральной нервной системы (околопроводное серое вещество, ядра шва ствола мозга, ядра ретикулярной формации, ядра таламуса, внутренней капсулы, мозжечка, интернейроны задних рогов спинного мозга и др.), оказывающих нисходящее тормозное действие на передачу ноцицептивной афферентации в дорзальных рогах спинного мозга. Основные нейроны антиноцецептивной системы локализованы в околоводопроводном сером веществе (сильвиев водопровод соединяет III и IV желудочки). Их аксоны образуют нисходящие пути к продолговатому и спинному мозгу и восходящие пути к ретикулярной формации, таламусу, гипоталамусу, лимбической системе, базальным ганглиям и коре. Медиаторами этих нейронов являются пентапептиды: метэнкефалин и лейэнкефалин. Энкефалины возбуждают опиатные рецепторы. Опиатные рецепторы возбуждаются не только медиаторами-энкефалинами, но и другими компонентами антиноцецептивной системы – гормонами головного мозга - эндорфинами (бета-эндорфин, динорфин). В механизмах развития анальгезии наибольшее значение придаётся серотонинергической, норадренергической, ГАМКергической и опиоидергической системам мозга. Основная из них, опиоидергическая система, образована нейронами, тело и отростки которых содержат опиоидные пептиды (бета-эндорфин, мет-энкефалин, лей-энкефалин, динорфин). Связываясь с определёнными группами специфических опиоидных рецепторов (мю-, дельта- и каппа-опиоидные рецепторы), 90% которых расположено в дорзальных рогах спинного мозга, они способствуют высвобождению различных химических веществ (гамма-аминомасляная кислота), тормозящих передачу болевых импульсов. Энкефалины и эндорфины возбуждают опиатные рецепторы. В энкефалинергических синапсах опиатные рецепторы находятся на постсинаптической мембране, но эта же мембрана является пресинаптической для других синапсов. Опиатные рецепторы ассоциированы с аденилатциклазой и вызывают ее ингибирование, нарушая в нейронах синтез цАМФ. В итоге уменьшается вход кальция и освобождение медиаторов, включая медиаторы боли (субстанция P, холецистокинин, соматостатин, глутаминовая кислота).
К медиаторам антиноцецептивной системы относятся также катехоламины. Они возбуждают тормозные a2-адренорецепторы, осуществляя тем самым постсинаптическое торможение боли.
Виды клеточного торможения · Пресинаптическое направленно на торможение выделения медиатора из-за гиперполяризации всего нейрона. · Постсинаптическое – гиперполяризация следующего нейрона.
Говоря об антиноцицептивной системе, первым компонентом следует ставить: 1. Желатинозную субстанцию спинного мозга (в чувствительных ядрах тригеминуса видимо, есть нечто подобное). 2. Нисходящие гипоталамо-спинальные пути (возможность обезболивания путем гипноза, внушения и самовнушения). С аксонов в спинном мозгу или на ядрах тригеминуса тоже выделяются тормозные медиаторы.
Естественная болеутоляющая система так же важна для нормальной жизнедеятельности, как и болесигнализирующая система. Благодаря ей, незначительные повреждения типа ушиба пальца или растяжения связок вызывают сильные болевые ощущения только на короткое время - от несколько минут до нескольких часов, не заставляя нас страдать в течение дней и недель, что случилось бы в условиях сохранения боли до полного заживления.
Таким образом, физиологическая ноцицепция включает четыре основных процесса: 1. Трансдукцию - процесс, при котором повреждающее воздействие трансформируется в виде электрической активности в свободных неинкапсулированных нервных окончаниях (ноцицепторах). Их активация происходит либо вследствие прямых механических или термических стимулов, либо под воздействием эндогенных тканевых и плазменных алгогенов, образующихся при травме или воспалении (гистамин, серотонин, простагландины, простациклины, цитокины, ионы К+ и Н+ , брадикинин). 2. Трансмиссию - проведение возникших импульсов по системе чувствительных нервных волокон и путей в центральную нервную систему (тонкие миелиновые А-дельта и тонкие безмиелиновые С-афференты в составе аксонов спинномозговых ганглиев и задних спинномозговых корешков, спиноталамические, спиномезенцефалические и спиноретикулярные пути, идущие от нейронов задних рогов спинного мозга к образованиям таламуса и лимбико-ретикулярного комплекса, таламокортикальные пути к соматосенсорным и фронтальной зонам коры головного мозга). 3. Модуляцию - процесс изменения ноцицептивной информации нисходящими, антиноцицептивными влияниями центральной нервной системы, мишенью которых являются преимущественно нейроны задних рогов спинного мозга (опиоидергические и моноаминовые нейрохимические антиноцицептивные системы и система воротного контроля). 4. Перцепцию - субъективное эмоциональное ощущение, воспринимаемое как боль и формирующееся под воздействием фоновых генетически детерминированных свойств центральной нервной системы и ситуационно меняющихся раздражений с периферии.
23. - Экстремальные состояния. Отличия обморака, колапса, шока и комы. общий патогенез шока.
Экстремальные состояния - состояния, сопровождающиеся грубыми расстройствами метаболизма и жизненно важных функций и представляющие непосредственную опасность для жизни. Экстремальные состояния часто связаны с действием сверхсильных патогенных факторов.
|
||
|
|
Последнее изменение этой страницы: 2016-06-19; просмотров: 1109; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.108 (0.008 с.) |