Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Глава 5 Характеристика отдельных свойств нервной системыСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
5.1. Сила нервной системы Понятие о свойстве силы нервной системы выдвинуто И. П. Павловым в 1922 г. При изучении условно‑рефлекторной деятельности у животных было выявлено, что чем больше интенсивность раздражителя или чем чаще он применяется, тем больше ответная условно‑рефлекторная реакция. Однако при достижении определенной интенсивности или частоты раздражения условно‑рефлекторный ответ начинает снижаться. В целом эта зависимость была сформулирована как «закон силы» (рис. 5.1). Было отмечено, что у животных этот закон проявляется по‑разному: запредельное торможение, при котором начинается снижение условно‑рефлекторного ответа, у одних наступает при меньшей интенсивности или частоте раздражения, чем у других. Первые были отнесены к «слабому типу» нервной системы, вторые – к «сильному типу». Возникли и два способа диагностики силы нервной системы: по максимальной интенсивности однократного раздражения, еще не приводящего к снижению условно‑рефлекторной реакции (измерение силы через «верхний порог»), и по наибольшему числу раздражения, тоже еще не приводящему к снижению рефлекторного ответа (измерение силы через ее «выносливость»). В лаборатории Б. М. Теплова была обнаружена большая чувствительность лиц со слабой нервной системой по сравнению с теми, у кого та оказалась сильной. Отсюда возник еще один способ измерения силы: через быстроту реагирования человека на сигналы разной интенсивности. Субъекты со слабой нервной системой из‑за своей более высокой чувствительности реагируют на слабые и средние по силе сигналы быстрее, чем субъекты с сильной нервной системой. По сути, в этом случае сила нервной системы определяется по «нижнему порогу».
Рис. 5.1. Схема, показывающая проявление «закона силы». По вертикали – величина реакции; по горизонтали – сила разрушения. В том же исследовательском коллективе сила нервной системы стала определяться и по уровню активации ЭЭГ. Однако этот метод технически сложен для массовых обследований. До недавнего времени все эти способы измерения силы нервной системы не имели единого теоретического обоснования и поэтому рассматривались как независимые друг от друга, обнаруживающие различные проявления силы нервной системы, связанные, как казалось, с разными физиологическими механизмами. Поэтому оправданным было и требование изучать типологические проявления свойств сразу несколькими методиками, о чем говорилось в предыдущем параграфе. Однако возможно единое объяснение различных проявлений силы нервной системы (Е. П. Ильин, 1979), которое делает равноправными различные методики, с помощью которых устанавливается сила нервных процессов. Объединяющим фактором оказался уровень активации в покое (суждение о котором выносилось на основании уровня энерготрат в покое – рис. 5.2): у одних людей он выше, а у других ниже. Отсюда и различия в проявлении «закона силы».
Рис. 5.2. Распределение испытуемых с различными энерготратами в покое (уровнем активации) в группах с различной силой нервной системы. По вертикали – число лиц, 5; по горизонтали – уровень энерготрат (ккал/кг/ч): I – от 0.50 до 0,99; II – от 1,00 до 1,50; III – от 1,51 до 2,00; IV – от 2, 01 и выше. А – лица с малой силой нервной системы; Б – лица со средней силой нервной системы; В – лица с большой силой нервной системы. Сила нервной системы как реактивность. Для возникновения видимой ответной реакции (ощущение раздражителя или движение рукой) нужно, чтобы раздражитель превысил определенную (пороговую) величину или по крайней мере ее достиг. Это значит, что данный раздражитель вызывает такие физиологические и физико‑химические изменения раздражаемого субстрата, которые достаточны для появления ощущения или ответной двигательной реакции. Следовательно, чтобы получить ответную реакцию, надо достичь порогового уровня активации нервной системы. Но в состоянии физиологического покоя последняя уже находится на определенном уровне активированности, правда, ниже порогового. У субъектов со слабой нервной системой уровень активации в покое выше (это следует из того, что в покое у них выше потребление кислорода и энерготраты на 1 кг веса тела); соответственно они ближе к пороговому уровню активации, с которого начинается реагирование (рис. 5.3), чем лица с сильной нервной системой. Для доведения этого уровня до порогового им, как следует из схемы, нужен меньший по интенсивности раздражитель. Субъектам же с сильной нервной системой, у которых уровень активации покоя ниже, требуется большая величина раздражителя, чтобы довести уровень активации до порогового. Этим и обусловлены различия между «слабыми» и «сильными» по нижнему порогу раздражения (r 1 < r 2). При нарастании интенсивности одиночных раздражителей уровень активации (возбуждения) и величина (или быстрота, как при измерении времени реакции) реагирования повышаются. Однако субъекты со слабой нервной системой, начав реагировать раньше, чем лица с сильной нервной системой, раньше достигают и предельного уровня активации, при котором наблюдаются наибольшие и самые быстрые ответные реакции. После этого у них эффект реагирования снижается, в то время как у субъектов с сильной нервной системой он еще возрастает. Они достигают предела активации позже, при большей силе одиночного раздражителя (R 1 < R 2). Следовательно, и «верхний» порог у «слабых» меньший, чем у «сильных», т. е. запредельное торможение у первых наступает раньше, чем у вторых, при меньшей интенсивности достаточно сильного раздражителя (рис. 5.3).
Рис. 5.3. Схема, показывающая различия в силе нервной системы в зависимости от интенсивности раздражителя. По вертикали – уровень активации: а 1 – в состоянии покоя у лиц со слабой нервной системой; а 2 – у лиц с сильной нервной системой; нижняя сплошная линия – пороговый уровень активации покоя, с которого начинается реакция на раздражитель; верхняя сплошная линия – предельный уровень реагирования (А 1 – для лиц со слабой нервной системой; А 2 – для лиц с сильной нервной системой). По горизонтали – интенсивность раздражителя: r1 – нижний порог для лиц со слабой нервной системой, r2 – то же для лиц с сильной нервной системой; R1 – верхний порог раздражителя для лиц со слабой нервной системой, R2 – то же для лиц с сильной нервной системой; h1 – величина дополнительной активации, необходимая для достижения порога реагирования лицами со слабой нервной системой, h2 – то же для лиц с сильной нервной системой. На выявление этих различий в реагировании людей на стимулы разной интенсивности направлена методика, разработанная В. Д. Небылицыным и получившая краткое название «наклон кривой» (рис. 5.4; описание методики см. в приложении). В. Д. Небылицын выдвинул гипотезу, что диапазон между нижним (r) и верхним (R) порогами должен сохраняться от индивида к индивиду неизменным:
... R: r = сопst.
Автор исходил из того факта, что оба этих порога у лиц с сильной нервной системой выше, чем у лиц со слабой нервной системой.
Рис. 5.4. Изменение времени реакции на различные по интенсивности звуковые сигналы у лиц с сильной и слабой нервной системой. По вертикали – время реакции, мс; по горизонтали – громкость звука, дБ. Сплошная линия – данные для лиц с сильной нервной системой; штрихпунктирная – для лиц со слабой нервной системой. Пунктиром обозначена зона слабых и средних интенсивностей звука, используемых в методике В. Д. Небылицына. Из приведенной формулы следует, что и сильная и слабая нервная система должны выдерживать одну и ту же величину градиента (прироста) сверхпорогового раздражителя. Если принять абсолютный порог за нулевую точку отсчета величины физиологической силы раздражителя, то при увеличении его силы и сильная и слабая нервная система будут реагировать одинаково: увеличится в два раза сила раздражителя – во столько же раз возрастет и величина ответной реакции со стороны как сильной, так и слабой нервной системы. Из этого должно также вытекать, что различий между последними при выравнивании физиологической силы раздражителя не будет; в обеих нервных системах запредельное торможение возникнет при одной и той же физиологической силе раздражителя. Это означает, что ход кривой реагирования на стимулы разной физиологической силы сильной и слабой нервной системы совпадет. Таким образом, согласно данной гипотезе В. Д. Небылицына, различия в силе нервной системы обнаруживаются потому, что используется физическая шкала интенсивности раздражителя, в которой одинаковая физическая величина последнего является разной физиологической силой для сильной и слабой нервной системы. Причиной этого, как теперь стало ясно, служит их различная фоновая активированность: чем она выше, тем большим по физиологической силе становится физический раздражитель. Однако эта правдоподобная гипотеза В. Д. Небылицына остается пока не доказанной на практике. Более того, П. О. Макаров (1955) использовал разницу между верхним и нижним порогами в качестве показателя силы нервной системы: чем больше диапазон между порогами (который автором принят за энергетический потенциал), тем больше сила нервной системы. Но и эта гипотеза осталась непроверенной экспериментально. Сила нервной системы как выносливость. Многократное повторное предъявление одинакового по силе раздражителя через короткие интервалы времени вызывает явление суммации, т. е. усиление рефлекторных реакций за счет роста фоновой активации, так как каждое предыдущее возбуждение оставляет после себя след, и поэтому каждая последующая реакция испытуемого начинается на более высоком функциональном уровне, чем предыдущая (заштрихованная область на рис. 5.5).
Рис. 5.5. Схема, показывающая различия в силе нервной системы в зависимости от длительности действия раздражителя. По вертикали – уровень активации (обозначения те же, что на рис. 5.3). По горизонтали – интенсивность раздражителя (ось Б) и длительность действия раздражителя (ось Т) с постоянной интенсивностью R2. Заштрихована область суммации следов возбуждения (повышение уровня подпороговой активности). t1 – время действия на слабую нервную систему раздражителя R2, приводящее к достижению предела реагирования; t2 – то же для сильной нервной системы. Поскольку исходный уровень активации у субъектов со слабой нервной системой выше, чем у субъектов с сильной нервной системой, суммация возбуждения и связанное с ней возрастание реагирования (несмотря на постоянную по физическим параметрам силу раздражителя) у них быстрее достигнет предела, и быстрее наступит «тормозный» эффект, т. е. снижение эффективности реагирования. У лиц с сильной нервной системой из‑за более низкой активации покоя имеется больший «запас прочности», и поэтому суммация у них может продолжаться дольше без достижения предела реагирования. Кроме того, возможно, что и последний у «сильных» находится на более высоком уровне, чем у «слабых». (Это не нашло отражения на схеме, где гипотетически пределы реагирования для «сильных» и «слабых» обозначены одинаково; единственно, что не укладывается в эту схему – случай, когда у «слабых» предел реагирования будет большим, чем у «сильных».) Поскольку величина суммации возбуждения определяется длительностью действия раздражителя (временем (t) или количеством повторений раздражения (n)), сильная нервная система оказывается более выносливой. Это значит, что при многократных предъявлениях сигналов (внешних или внутренних – самоприказов) снижение эффекта реагирования на таковые (величины или быстроты реакций) у «слабых» произойдет быстрее, чем у «сильных». На этом и основаны различные методики определения силы нервной системы через ее выносливость. Следует обратить внимание на два существенных момента. Во‑первых, при диагностике силы нервной системы нельзя использовать слабые раздражители, так как они снижают, а не повышают активацию нервной системы, и в результате более выносливыми к монотонному раздражителю оказываются лица со слабой нервной системой. Кстати, по этому поводу еще в лаборатории И. П. Павлова возник спор: ее глава считал, что те собаки, которые быстро засыпали в «башне молчания» при выработке у них условных рефлексов, имеют слабую нервную систему. Однако его ученица К. П. Петрова (1934) доказала, что это как раз собаки с сильной нервной системой, не выдерживающие монотонной обстановки (или как сейчас сказали бы – сенсорной депривации). В конце концов И. П. Павлов признал правоту ученицы. Во‑вторых, не каждый показатель выносливости может служить критерием силы нервной системы. Выносливость к физической или умственной работе не является прямым индикатором силы нервной системы, хотя и связана с ней. Речь должна идти о выносливости именно нервных клеток, а не человека. Поэтому методики должны показывать быстроту развития запредельного торможения, с одной стороны, и выраженность эффекта суммации – с другой. 5.2. Подвижность – инертность и лабильность нервных процессов Выделенное И. П. Павловым в 1932 г. свойство подвижности нервных процессов стало в дальнейшем, как отмечал Б. М. Теплов (1963а), оцениваться как более многозначное. Поэтому он выделил следующие особенности нервной деятельности, характеризующие быстроту функционирования нервной системы: 1) быстроту возникновения нервного процесса; 2) быстроту движения нервного процесса (иррадиация и концентрация); 3) быстроту исчезновения нервного процесса; 4) быстроту смены одного нервного процесса другим; 5) быстроту образования условного рефлекса; 6) легкость переделки сигнального значения условных раздражителей и стереотипов. Изучение взаимосвязи между этими проявлениями быстроты функционирования нервной системы, проведенное в лаборатории Б. М. Теплова, позволило выделить два главных фактора: легкость переделки значения условных раздражителей (положительного на отрицательный и наоборот) и быстроту возникновения и исчезновения нервных процессов. За первым фактором Б. М. Теплов оставил название подвижность, а второй обозначил как лабильность. Другие показатели быстроты функционирования нервной системы не относятся в настоящее время к двум указанным свойствам. Попытка М. Н. Борисовой выделить быстроту иррадиации и концентрации нервных процессов в самостоятельное свойство не получила достаточно веских аргументов. Также неудачной, как уже говорилось, оказалась попытка В. Д. Небылицына выделить быстроту образования условных рефлексов в отдельное свойство динамичности. Хотя переделка до сих пор используется в ряде физиологических работ как показатель подвижности нервной системы, полученные в последние десятилетия данные поставили ее как референтный показатель свойства подвижности под сомнение. Оказалось, что переделка условных рефлексов – довольно сложный феномен высшей нервной деятельности, который определяется не только легкостью перехода возбуждения в торможение и обратно, но и прочностью образованных условных связей (т. е. быстротой затухания следов), интенсивностью раздражителя, влиянием второй сигнальной системы и т. д. (В. А. Трошихин и др., 1978). Да и сам И. П. Павлов расценивал переделку условных раздражителей как весьма сложное комплексное испытание, довольно трудно поддающееся расшифровке. Переделка не связана с другими индикаторами подвижности, в частности с показателями, входящими в группу лабильности. Зато она обнаруживает зависимость от силы нервной системы. В связи с этим физиологическое толкование «переделки» как свойства нервной системы весьма затруднительно. По крайней мере, очевидно, что она не является простым аналогом быстроты течения нервных процессов. Поэтому не случайно в последние два десятилетия больше изучаются показатели группы лабильности, т. е. быстрота развития и исчезновения нервных процессов. Этому способствует и то обстоятельство, что «переделка» требует весьма продолжительного времени, поэтому к ней невозможно обращаться при массовых обследованиях. Исходя из того, что лабильность предполагает быстроту развития нервного процесса и быстроту его исчезновения, наметились три методических подхода в изучении функциональной подвижности (лабильности): 1) выявление быстроты возникновения возбуждения и торможения; 2) выявление быстроты исчезновения возбуждения и торможения; 3) выявление максимальной частоты генерации нервных импульсов, зависящей как от первого, так и от второго. Изучение быстроты развития нервных процессов значительно усложнено тем, что она зависит, как говорилось в предыдущем параграфе, от уровня активации покоя, т. е. от того, какова нервная система у обследуемого – слабая или сильная. Конечно, это не исключает влияния на быстроту генерации возбуждения и других механизмов, которые могут прямо характеризовать предполагаемое свойство нервной системы. Однако выделить их в «чистом» виде пока нельзя. Еще хуже обстоит дело с измерением быстроты возникновения торможения. Сейчас можно рассчитывать лишь на один способ – измерение латентного периода расслабления мышц с помощью электромиографии. Функциональная подвижность как быстрота исчезновения нервных процессов. Нервный процесс не исчезает сразу после действия раздражителя или осуществления какого‑то действия, а ослабевает постепенно. Наличие следов препятствует нормальному развитию противоположного нервного процесса. Однако, даже исчезнув, первый процесс не перестает влиять на развитие ему противоположного. Дело в том, что по механизму индукции он сменяется фазой, облегчающей возникновение такового. Например, вместо бывшего раньше процесса возбуждения возникает в этих же центрах процесс торможения. Если на этом фоне воздействовать тормозным раздражителем, возникшее торможение суммируется с уже имевшимся индукционным торможением и тогда тормозной эффект усиливается. Временная развертка происходящих изменений представлена на рис. 5.6. Последействие, зависящее от следовой деполяризации и циркуляции нервных импульсов по сети нейронов, у разных людей имеет разную длительность. У одних положительные и отрицательные фазы протекают быстро, у других – медленно. Поэтому, если предъявлять разным людям одинаковые задачи на сближение положительных и тормозных раздражителей или возбудительных и тормозных реакций, выявляются разные временные развертки происходящих следовых изменений, т. е. различия по функциональной подвижности нервной системы.
Рис. 5.6. Схема, показывающая фазность развития следовых процессов. А – изменение величины тормозных реакций после предшествования процесса возбуждения; Б – изменение величины активизирующих реакций после предшествования тормозных реакций. Столбиками обозначена величина реакций, кривые линии – изменение во времени нервных процессов (t0–t5): следовое возбуждение, а1 – исчезновение следов возбуждения, а2–а4 – торможение, развивающееся по механизму отрицательной индукции; б0 – следовое торможение, б1 – исчезновение следового торможения, б2–б5 – возбуждение, развивающееся по типу положительной индукции. Поскольку длительность затухания следов нервных процессов может зависеть от их интенсивности (чем интенсивнее процесс, тем длительнее будет его затухание), важно учитывать влияние этого фактора. У лиц со слабой нервной системой при действии одного и того же раздражителя процесс возбуждения развивается более интенсивно (по крайней мере – в пределах слабых и средних по интенсивности раздражителей), затухание же его будет более длительным, чем у людей с сильной нервной системой. Не случайно в психофизиологической лаборатории Б. М. Теплова – В. Д. Небылицына выявлены положительные связи между инертностью и слабостью нервной системы. Однако при нивелировании разными методическими приемами различий в уровне активации покоя можно получить показатель быстроты протекания следовых процессов в чистом виде. Так, не найдены корреляции между силой нервной системы и подвижностью нервных процессов при использовании для выявления последействия методик К. М. Гуревича и Е. П. Ильина, о которых речь будет идти ниже (см. приложение). Методики, изучающие функциональную подвижность по быстроте течения следовых явлений, чаще всего основаны на том, что после положительного сигнала, иницирующего возбудительный процесс, предъявляется тормозной сигнал, вызывающий противоположный процесс или реакцию. И наоборот, после тормозного сигнала (или реакции) через короткое время предъявляется положительный сигнал, вызывающий возбудительную реакцию. Эти приемы весьма близки к приему, названному И. П. Павловым «сшибкой». Однако они не тождественны приему, названному «переделкой» сигнального значения раздражителей, хотя в том и другом случае есть внешне сходный момент: один нервный процесс (или реакция) должен уступить место другому. Различие этих двух приемов, как отмечают В. А. Трошихин и его соавторы, состоит в следующем. При «сшибке» смена одного нервного процесса другим обусловлена последовательным действием двух различных сигналов или операций (например, звука как положительного раздражителя и света как отрицательного). При «переделке» же меняется сигнальное значение одного и того условного раздражителя, остающегося неизменным по своей модальности и физическим параметрам. При «сшибке» имеется столкновение в один и тот же момент времени двух процессов, в «переделке» – разновременная смена положительного и тормозного раздражителей. «Переделка» связана с угашением упроченной условно‑рефлекторной реакции и выработкой на тот же раздражитель условного тормоза.
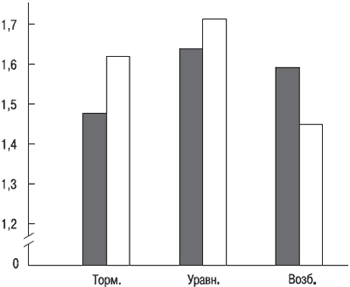
5.3. Баланс нервных процессов Соотношение нервных процессов было первым из свойств нервной системы, указанных И. П. Павловым. Несмотря на это, оно до сих пор наименее изучено. Во всяком случае, мы не можем говорить о том, что исследуем баланс нервных процессов так, как его понимал И. П. Павлов (напомним, что он говорил о балансе по силе возбуждения и силе торможения). Не можем так заявлять, поскольку не знаем способов определения силы процесса торможения. Вместо этого мы судим (по косвенным признакам) о превалировании или равновесии возбудительных и тормозных реакций в действиях человека. В качестве показателей этого свойства у разных исследователей павловской школы выступали: величина положительных и тормозных условно‑рефлекторных реакций, отношение числа ошибок (или же правильных реакций) на положительный и тормозной сигнал, постоянство фона условно‑рефлекторной деятельности и др. (Э. П. Кокорина, 1963; Г. А. Образцова, 1964 и др.). В психологии при измерении баланса нервных процессов у человека используются другие показатели: число переводов и недоводов при воспроизведении на основе проприорецепции (при выключении зрения) амплитуды движений, а также временных отрезков (Г. И. Борягин, 1959; М. Ф. Пономарев, 1960, и др.). По мнению этих исследователей, наличие переводов свидетельствует о преобладании возбуждения, а наличие недоводов – о преобладании торможения. Эти представления находят подтверждение как в опытах с фармакологическими воздействиями на человека, так и в исследованиях, проводимых на различном эмоциональном фоне испытуемого. Так, прием обследуемыми кофеина, усиливающего возбуждение, приводит к увеличению срывов дифференцировок (по которым судят о выраженности торможения) и возрастанию количества переводов при воспроизведении амплитуд движений. Прием же брома, усиливающего тормозной процесс, уменьшает количество срывов дифференцировок и увеличивает число недоводов при воспроизведении амплитуд (Г. И. Борягин, М. Ф. Пономарев). В состоянии предстартового возбуждения, регистрируемого как по самоотчету спортсменов, так и по ряду физиологических показателей (пульсу, артериальному давлению, тремору и т. п.), резко увеличивается число переводов воспроизводимых амплитуд движений, а в состоянии заторможенности (при скуке, сонливости) возрастает число недоводов. Однако все это говорит о соотношении возбуждения и торможения по их величине (интенсивности), но не по силе в смысле выносливости нервной системы, как понимал баланс И. П. Павлов. Однако так сложилось, что всегда имелся в виду баланс именно в павловском его толковании, и никто не обращал внимания на то, что легче всего (и ближе к истине) говорить о соотношении величины возбуждения и торможения и изучать влияние именно этого соотношения на поведение и деятельность человека. По крайней мере, имеющиеся у физиологов и психологов методы изучения баланса нервных процессов не дают возможности рассчитывать на большее. Особенностью изучения баланса между возбуждением и торможением по их величине является то, что о нем судят по интегральной характеристике, результирующей противоборства этих двух процессов (или систем реагирования – возбудительной и тормозной). Таким образом, у разных людей сравниваются не выраженность возбуждения или торможения, а какой из процессов берет верх над другим. Поэтому теоретически одна и та же типологическая особенность у двух субъектов (например, преобладание возбуждения над торможением) может базироваться на разных уровнях выраженности того и другого. Так, у одного субъекта преобладание возбуждения над торможением происходит при высокой интенсивности обоих, а у второго преобладание возбуждения может наблюдаться при слабой их выраженности. Попытка глубже разобраться в физиологической сущности этого свойства привела к выявлению ряда интересных фактов, которые, однако, окончательного ответа не дают. Например, обнаружено, что баланс, как и сила нервной системы, связан с уровнем активации покоя (Е. П. Ильин, 1999). Однако если для силы нервной системы такая связь имеет линейный характер (чем слабее нервная система, тем выше активация в покое), то для баланса – криволинейный: уровень активации (энерготраты в покое на 1 кг веса человека) выше у лиц с уравновешенностью возбуждения и торможения и ниже у лиц с преобладанием возбуждения и торможения (см. рис. 5.7).
Рис. 5.7. Энерготраты в состоянии физиологического покоя у субъектов с различными типологическими особенностями по балансу нервных процессов. По вертикали – энерготраты (кал/кг/ч); по горизонтали – типологические особенности по балансу. Заштрихованные столбики – «внешний» баланс, незаштрихованные – «внутренний» баланс. Такая криволинейная связь баланса с уровнем активации покоя подтверждается наличием криволинейной связи баланса с силой нервной системы: слабости нервной системы чаще соответствует уравновешенность нервных процессов, а силе – неуравновешенность (преобладание возбуждения или торможения). В соответствии с обнаруженной связью у уравновешенных уровень активации покоя в среднем должен быть выше, чем у неуравновешенных (поскольку уровень активации выше у «слабых»). Однако обращает на себя внимание одно обстоятельство: средний уровень активации покоя у уравновешенных ниже такого же показателя у слабых (вероятно, потому что не все уравновешенные имеют слабую нервную систему, т. е. наибольший уровень активации покоя). Эти факты, хотя и не дают прямого ответа на вопрос о физиологической природе изучаемого свойства нервной системы, наталкивают на мысль, что при рассмотрении соотношения между возбуждением и торможением следует, очевидно, отказаться от, казалось бы, простой и очевидной схемы: баланс – это прямая линия, на верхнем конце которой господствует возбуждение, а на нижнем – торможение; уравновешенность же – срединная точка на этой линии, свидетельствующая о средней выраженности того и другого процесса. Полученные данные не укладываются в подобную схему: преобладание возбуждения и преобладание торможения – не два полюса одной прямой, а отношения между ними гораздо сложнее, и уравновешенность не является промежуточной (срединной) инстанцией между ними. Это предположение подкрепляется и другими фактами. Первый состоит в том, что при измерении «внешнего» баланса в середине ночи сразу после пробуждения обследуемых было выявлено: «возбудимые» и «тормозные» по дневным измерениям ночью переходили в разряд уравновешенных. Если переход первых в уравновешенные удивления не вызывал и соответствовал представлениям об усилении тормозных процессов во время сна, то переход «тормозных», что должно расцениваться как усиление возбуждения, никак не укладывался в общепринятые представления. Правда, такой переход наблюдался не у всех испытуемых, но все же показатели 9 из 17 «возбудимых» и 12 из 17 «тормозных», что ночью перешли в разряд уравновешенных (Е. П. Ильин и М. И. Семенов, 1969). Обратил на себя внимание и тот факт, что ночью повысилась точность воспроизведения, как будто бы испытуемым стало легче выполнять задание экспериментатора. Этот факт навел на мысль, что в полудремотном состоянии люди освобождались от давившего на них днем и мешавшего действовать раскрепощенно мотивационного фактора. Наблюдение за поведением испытуемых во время ночного эксперимента, когда у них было одно желание – скорее отделаться от экспериментатора и продолжить сон, позволило предложить: и худшая точность воспроизведения, и частое появление случаев с преобладанием возбуждения или торможения при дневных измерениях могли быть следствием желания испытуемого выполнить задание экспериментатора как можно лучше. Ночью же это «давление» на двигательные действия обследуемых либо исчезало, либо значительно ослаблялось, отсюда в том и другом случае управление движениями было разным. Еще в одном исследовании вмешательство стремления «сделать лучше» в управление точностными движениями было устранено благодаря гипнозу (Е. П. Ильин, С. К. Малиновский, 1981). Испытуемые, у которых баланс измерялся в состоянии бодрствования, вводились в первую стадию гипноза, во время которой они выполняли под команду экспериментатора ту же самую пробу для определения баланса. Из 16 человек у 3 было в обычном состоянии преобладание возбуждения, и именно их не удалось перевести в гипнотическое состояние и выяснить, достигнут ли они уравновешенности. Однако для нас важнее было выяснить, придут ли к последней лица с преобладанием торможения (таких мы подобрали 6 человек). Наши ожидания подтвердились: 5 из 6 испытуемых в состоянии гипнотического сна перешли в уравновешенные. Таким образом, подтвердились результаты эксперимента с прерыванием естественного ночного сна. А это значит, что в полусонном состоянии наши испытуемые освобождались как от тормозных, так и от возбудительных влияний на управление движениями на точность в пространстве. Чем вызваны эти влияния, можно только предполагать (скорее всего они проистекают из лобных отделов больших полушарий, в которых находятся интегративные центры, заведующие сознательными актами человека). При блокировании во время сна таких влияний центры управления движениями переходят на автоматизированный и более оптимальный режим. Соответственно можно предположить, что уравновешенность нервных процессов является исходной базовой характеристикой при автоматизированном режиме работы нервных центров, а преобладание возбуждения или торможения – искажение этого соотношения нервных процессов в результате вмешательства другого уровня управления, связанного с активным привлечением внимания человека к выполняемому заданию, с его желанием выполнить таковое как можно лучше. Какое соотношение между возбуждением и торможением проявится у данного человека, зависит, вероятно, от его типа реагирования на ситуацию: у одних типичная – возбудительная реакция, у других – тормозная, у третьих же возникает индифферентная реакция или вообще таковая отсутствует, поэтому у них проявляется базовое соотношение между возбуждением и торможением, т. е. их уравновешенность. Несмотря на то что данное объяснение природы баланса не больше чем гипотеза, только оно позволяет на данном уровне наших знаний как‑то объяснить те факты с изменениями баланса и его связь с уровнем активации покоя, которые были выявлены. Ясно только одно: сущность свойства уравновешенности по величине возбудительных и тормозных реакций нуждается в дальнейшем изучении, и на этом пути нас ожидает, вероятно, еще много неожиданного. Есть основания полагать, что баланс между величиной возбуждения и торможения на разных контурах регулирования центральной нервной системы выражен неодинаково. Так, кроме баланса, о котором речь шла выше и названного «внешним», существует и другой вид баланса, названный «внутренним». Он получил такое название потому, что, с одной стороны, не реагирует на изменение эмоционального состояния человека, например на предстартовое возбуждение; с другой – отражает уровень активации, связанный с потребностью в двигательной активности, т. е. этот баланс связан с более глубинными (внутренними) процессами в центральной нервной системе. Нетождественность «внешнего» и «внутреннего» баланса показывает ряд фактов. Во‑первых, между ними нет прямых корреляций (ни положительных, ни отрицательных). Во‑вторых, при ряде состояний человека (монотонии, психическом пресыщении) сдвиги по этим балансам бывают разнонаправленными: сдвигу «внешнего» баланса в сторону возбуждения соответствует сдвиг «внутреннего» в сторону торможения, а сдвигу «внешнего» баланса в сторону торможения соответствует сдвиг «внутреннего» в сторону возбуждения. Это связано с механизмами саморегуляции уровня активации в центральной нервной системе, «переливанием» активности из одного уровня регуляции в другой (А. А. Крауклис, 1963). В‑третьих, у «внешнего» и «внутреннего» баланса имеются свои специфичные проявления в особенностях поведения и деятельности спортсменов, что находит отражение и в том, сколь часто встречаются типологические особенности проявления этих свойств у представителей различных видов спорта. Например, если преобладание возбуждения по «внешнему» балансу более типично для спортсменов, специализирующихся в «коротком» спринте, то преобладание возбуждения по «внутреннему» балансу присуще спортсменам, предпочитающим «длинный» спринт, который требует скоростной выносливости. Возможно, в этих двух видах баланса проявляют себя две системы активации – ретикулярная формация и гипоталамус. Однако само наличие этих систем как самостоятельных оспаривается некоторыми физиологами. «Внутренний» баланс также связан криволинейной зависимостью с уровнем активации покоя: наибольший уровень таковой наблюдается у лиц с уравновешенностью (однако он ниже, чем у лиц с уравновешенностью по «внешнему» балансу). Большинство методик изучения баланса возбуждения и торможения (рефлексометрическая методика Ю. А. Поворинского, кожно‑гальваническая методика В. С. Мерлина и др.) основаны на выработке положительных и тормозных реакций и учитывают количество сорванных дифференцировок (т. е. реакций на тормозной раздражитель). Критерии деления на уравновешенных и неуравновешенных хотя и количественные, но весьма условные, а главное, дающие возможность судить о сдвиге баланса только в одну сторону – возбуждения. Сдвиг же баланса в сторону торможения не учитывается. В связи с этим данные методики не позволяют проводить полноценную диагностику типологических особенностей по «внешнему» бал
|
||||
|
|
Последнее изменение этой страницы: 2016-04-23; просмотров: 593; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.133.153.224 (0.016 с.) |