Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Взаимодействие системы терморегуляции с другими физиологическими системами организмаСодержание книги
Поиск на нашем сайте
Система терморегуляции использует для осуществления своих функций эффекторные компоненты других физиологических систем. Это обстоятельство обусловливает необходимость постоянного взаимодействия, сопряжения или конкуренции механизмов, регулирующих теплообмен и другие гомеостатические функции. Такое сопряжение регуляции теплообмена и других гомеостатических функций имеет место, прежде всего, в гипоталамусе, где термочувствительные нейроны преоптической области гипоталамуса являются одновременно чувствительными к изменению осмотического давления, артериального давления крови, концентрации ионов Н+, Na+, Са2+, СО2, глюкозы. Эти нейроны изменяют свою биоэлектрическую активность при сдвигах температуры тела, под действием эндопирогенов, половых гормонов, некоторых нейромедиаторов. Таким образом, центр терморегуляции в гипоталамусе постоянно взаимодействует с другими расположенными здесь же центрами регуляции гомеостаза. Используемые организмом в системе регуляции теплообмена нейромедиаторы, гуморальные вещества также одновременно принимают участие в регуляции других функций и показателей гомеостаза. Их примерами могут быть катехоламины, которые параллельно выполняют функции медиаторов в центральной и симпатической нервных системах, функции сосудоактивных веществ, активаторов обменных процессов. 13.5.1. Сердечно-сосудистая система и терморегуляция В качестве эффекторов в реакциях теплообмена используются сосуды поверхности тела, посредством которых регулируется кровоток в коже, ее температура и интенсивность теплоотдачи. В термонейтральных условиях, при действии на организм умеренно низких температур или неглубокой гипотермии изменение кровотока в поверхностных тканях не оказывает существенного влияния на деятельность сердца и системную гемодинамику. При действии же на организм высоких температур, гипертермии, лихорадке резкое расширение сосудов поверхности тела, влияние высокой температуры на центральные механизмы регуляции кровообращения могут привести к падению давления крови, развитию коллаптоидного состояния. Использование при гипертермии многочисленных поверхностных сосудов, как общих эффекторов сердечно-сосудистой и терморегулирующей систем, соподчинено более важной в этот момент времени гомеостатической потребности организма — поддержанию системного кровотока.
13.5.2. Водно-солевой баланс и терморегуляция Когда температура поверхности тела достигает величин, равных температуре окружающей среды, ведущее значение в Механизмах теплоотдачи приобретает уже не повышение кровотока в поверхностных тканях, а потоотделение и испарение пота и влаги с поверхности тела. Более существенную роль начинают играть учащение дыхания и испарение влаги с поверхности дыхательных путей. Включение в реакции теплообмена потоотделения является примером использования общих эффекторов для системы терморегуляции и систем регуляции водно-солевого баланса и осмотического давления. Если при подъеме температуры тела в силу потери жидкости за счет усиленного потоотделения уменьшается объем циркулирующей крови и повышается ее осмотическое давление, организм стремится сохранить водный гомеостаз, даже если это идет в ущерб терморегуляторным реакциям. С развитием гипогидратации и повышением осмотического давления в организме отдача тепла за счет потоотделения уменьшается и температура тела устанавливается на еще более высоком уровне. Развивается чувство жажды, уменьшается диурез. В конкуренции за общие эффекторные механизмы начинают преобладать системы осмо- и волюморегуля- ции, как более древние и в экстремальных условиях более важные для сохранения гомеостаза. Сопряжение осмо- и терморегуляции достигается в нервных центрах медиальной преоптической области гипоталамуса, где тепло- и холодочувствительные нейроны наделены одновременно высокой осмочувствительностью. Подтверждением сопряженного протекания в организме процессов термо- и осморегуляции являются изменения водного обмена противоположной направленности — при охлаждении организма. При действии на организм низкой температуры имеют место уменьшение потребления воды, усиление диуреза и повышение осмолярности плазмы крови. Если дегидратация при действии на организм высокой внешней температуры приводит к торможению терморегуляторных реакций, то при действии на него низкой температуры дегидратация ведет к торможению теплочувствительных нейронов гипоталамуса и в результате — к снижению теплоотдачи.
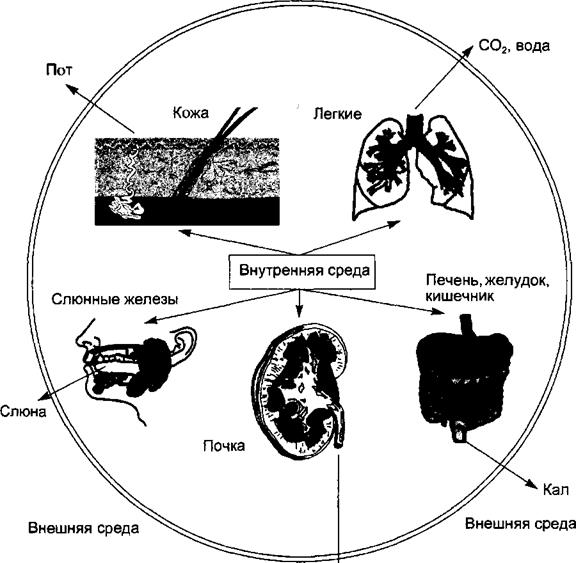
13.5.3. Дыхание и терморегуляция При действии на организм высокой внешней температуры активация потоотделения и дыхания ведет к усиленному выделению из организма СО2, некоторых минеральных ионов. Как при гипер-, так и при гипотермии могут наблюдаться сдвиги кислотно-основного состояния. За счет полипноэ и интенсификации потоотделения развивается дыхательный алкалоз, сопровождающиейся увеличением pH и снижением рСО2 в крови. При нарастании гипертермии в связи с ухудшением доставки к тканям кислорода в них развивается метаболический ацидоз. Смена щелочной реакции крови на кислую при выраженной гипертермии вновь начинает играть положительную регуляторную роль, как для усиления теплоотдачи, так и для предупреждения дальнейшего закисления крови и улучшения оксигенации тканей. Это достигается стимуляцией дыхательного центра посредством избытка Н2+, увеличения минутного объема дыхания, усиления испарения влаги с поверхности дыхательных путей, что ведет к снижению рСО2 и увеличению рО2. Обратные взаимоотношения между процессами регуляции теплообмена и дыхания прослеживаются при гипотермии. Развивающаяся при этом гиповентиляция является общим эффекторным механизмом, обеспечивающим снижение теплопотерь, поддержание на более низком уровне pH крови соответственно сниженной температуре тела. ГЛАВА 14 Выделение. Функции почек. Водно-солевой обмен Выделение — часть обмена веществ, осуществляемая путем выведения из внутренней среды организма во внешнюю среду конечных и промежуточных продуктов метаболизма, чужеродных и излишних веществ для обеспечения оптимального состава внутренней среды и нормальной жизнедеятельности организма. Процессы выделения являются неотъемлемым признаком жизни, поэтому их нарушение неизбежно приводит к нарушениям гомеостазиса, обмена веществ и функций организма, вплоть до его гибели. Выделение неразрывно связано с обменом воды, поскольку основная часть предназначенных для выведения из организма веществ выделяется в растворенном виде. Основным органом выделения являются почки, образующие и выделяющие мочу и вместе с ней вещества, подлежащие удалению из организма. Почки являются также основным органом обеспечения водно-солевого обмена, поэтому в этой главе и рассматриваются функции почек, выделение и водно-солевой обмен. Органы и процессы выделения Функция выделения веществ из внутренней среды организма осуществляется почками, желудочно-кишечным трактом, легкими, кожей и слизистыми оболочками, слюнными железами. Реализуемые ими процессы выделения находятся в координированной взаимосвязи и поэтому функционально эти органы объединяют понятием выделительная система организма (рис. 14.1). Между органами выделения существуют функциональные и регуляторные взаимосвязи, в результате чего сдвиг функционального состояния одного из органов выделения меняет активность другого в пределах единой выделительной системы. Так, например, при избыточном выведении жидкости через кожу путем потоотделения при высокой температуре —- снижается объем мочеобразования, при уменьшении экскреции азотистых соединений с мочой — увеличивается их выведение через желудочно-кишечный тракт, легкие и кожу.
14.1.1. Выделительная функция кожи Выделительная функция кожи, непосредственно контактирующей с внешней средой большой площадью поверхности, преимущественно обеспечивается деятельностью потовых и, в меньшей степени, сальных желез. В среднем у человека за сутки выделяется от 300 до 1000 мл пота, что зависит от температуры окружающей среды и интенсивности энергетического метаболизма. Составы пота и плазмы крови отличаются, поскольку пот является не простым фильтратом плазмы, а секретом потовых желез. С потом из организма выводится в покое до !/3 общего количества экскретируе- Внешняя среда
Моча Рис. 14.1. Выделительная система организма.
Органы выделения (кожа, легкие, слюнные железы, печень и желудочно-кишечный тракт, почки) выводят из внутренней среды организма во внешнюю среду метаболиты и ксенобиотики в составе секретов и экскретов. мой воды, 5—7 % всей мочевины, мочевая кислота, креатин, хлориды, натрий, калий, кальций, органические вещества, липиды, микроэлементы. Через кожу может выделяться даже больше кальция, чем выводится с мочой. При недостаточности функции почек или печени возрастает выделение через кожу веществ, обычно экскретируемых с мочой, — мочевины, ацетона, желчных пигментов и др. С потом выделяются пепсиноген, амилаза и щелочная фосфатаза, отражая тем самым функциональное состояние органов пищеварения. Регуляция потоотделения осуществляется симпатическими холинергическими влияниями, а также гормонами — вазопрессином, альдостероном, гормонами щитовидной железы и половыми стероидами. Секрет сальных желез на 2/3 состоит из воды, а г/3 составляют неомыляемые соединения — холестерин, сквален (алифатический углеводород), аналоги казеина, продукты обмена половых гормонов, кортикостероидов, витаминов и ферментов. В выделительной системе сальные железы не имеют большой значимости, так как за сутки выделяется всего лишь около 20 г секрета. Регуляция сальных желез осуществляется в основном половыми и надпочечниковыми стероидами. 14.1.2. Выделительная функция печени и пищеварительного тракта Выделительная функция печени реализуется за счет образования и секреции в ней желчи. За сутки печень секретирует от 500 до 2000 мл желчи, но большая часть ее объема затем реабсорбируется в желчном пузыре и кишечнике. С желчью из организма экскретируются конечные продукты обмена гемоглобина и других порфиринов в виде желчных пигментов, конечные продукты обмена холестерина — в виде желчных кислот. Несмотря на всасывание в кишечнике и обратный транспорт в печень с кровью воротной вены, часть этих веществ покидает организм с фекальными массами. В составе желчи из организма выделяются тироксин, мочевина, кальций и фосфор, а также вещества, поступающие в организм: лекарственные препараты, ядохимикаты и др. В желчном пузыре происходит обратное всасывание в кровь воды и растворенных в ней веществ, прежде всего электролитов. Этот процесс приводит к концентрированию желчи и регулируется гормоном вазопрессином, повышающим проницаемость стенки желчного пузыря.
Выделительная функция желудка обеспечивает выведение в составе желудочного сока продуктов метаболизма (мочевины, мочевой кислоты), лекарственных и ядовитых веществ (ртуть, йод, салицилаты, хинин). Выделительная функция кишечника состоит, во-первых, в выделении продуктов распада пищевых веществ, не подвергшихся всасыванию в кровь и представляющих излишние или вредные для организма соединения. Во-вторых,, кишечник экскретирует вещества, поступившие в его просвет с пищеварительными соками (желудочным, поджелудочным) и желчью. При этом многие из них в кишечнике подвергаются метаболизму и с калом выделяются не сами вещества, а их метаболиты, например метаболиты билирубина желчи. В-третьих, стенка кишечника способна экскретировать из крови ряд веществ, среди которых особое значение имеет экскреция плазменных белков. При чрезмерности этого процесса возникает избыточная потеря организмом белка, ведущая к патологии. Из крови кишечный эпителий экскретирует соли тяжелых металлов, магний, почти половину всего выделяемого организмом кальция. Вместе с экскрементами кишечником выделяется и некоторое количество воды (в среднем около 100 мл/сут). 14.1.3. Выделительная функция легких и верхних дыхательных путей Процессы газообмена, происходящие в легких, обеспечивают удаление из внутренней среды организма летучих метаболитов и экзогенных веществ — углекислого газа, аммиака, ацетона, этанола, метилмеркаптана и др. Кроме того, за счет мерцательного эпителия удаляются продукты обмена веществ самой легочной ткани и эпителия воздухоносных путей, например продукты деградации сурфактанта. Легкие выделяют в просвет дыхательных путей небольшие количества белка, в том числе гамма-глобулинов, обладающих сродством к легочной ткани, а также входящих в состав секрета желез бронхиального дерева. Через слизистую оболочку дыхательных путей испаряется значительное количество воды (от 400 мл в покое до 1 л при усиленном дыхании), а при повышении проницаемости аэрогематиче- ского барьера из крови могут в избытке выделяться пурины, аденозин- и гуанозинмонофосфаты. Гиперсекреция желез слизистой оболочки верхних дыхательных путей имеет место при нарушениях выделительной функции почек, в этом случае через слизистую оболочку выделяется много мочевины, которая, разлагаясь, образует аммиак, определяющий соответствующий запах изо рта.
Функции почек Функции почек многообразны, при этом часть из них связана с процессами выделения, в которых почки играют ведущую роль, другая же часть подразумевает невыделительные функции почек. Почки участвуют в регуляции: 1) водного баланса организма и, соответственно, объемов вне- и внутриклеточных водных пространств, поскольку меняют количество выводимой с мочой воды; 2) ионного баланса и состава жидкостей внутренней среды путем избирательного изменения экскреции ионов с мочой; 3) постоянства осмотического давления жидкостей внутренней среды за счет изменения количества выводимых осмотически активных веществ (солей, мочевины, глюкозы и др.); 4) кислотно-основного баланса, путем изменения экскреции водородных ионов, нелетучих кислот и оснований; 5) метаболизма белков, липидов, углеводов, нуклеиновых кислот и других органических соединений, во-первых, за счет изменений экскреции продуктов метаболизма и избытка соединений, поступивших с пищей или образовавшихся в организме, во-вторых, благодаря собственной метаболической функции (синтез аммиака и мочевины, новообразование глюкозы, гидролиз белков и липидов, синтез ферментов, простаноидов и т. п.); 6) циркуляторного гомеостазиса, путем регуляции обмена электролитов, объема циркулирующей крови, внутренней секреции гормонов, регулирующих функции сердечно-сосудистой системы, — ренина, кальцитриола и др., а также экскреции катехоламинов и других гормональных регуляторов системы кровообращения; 7) эритропоэза, за счет внутренней секреции эритропоэтина — гуморального регулятора эритрона; 8) гемостаза, путем образования гуморальных регуляторов свертывания крови и фибринолиза (урокиназы, тромбопластина, тромбоксана и простациклина) и участвуя в обмене физиологических антикоагулянтов (гепарина). Экскретируя из внутренней среды чужеродные и вредные вещества, почки выполняют защитную функцию. Таким образом, выделяют следующие функции почек: экскреторную, гомеостатическую, метаболическую, инкреторную и защитную. Основной функцией почек, обеспечивающей ведущую роль в выделительной системе организма, является образование и выделение мочи. 14.2.1. Механизмы мочеобразования Моча образуется в почках из плазмы крови, причем почка относится к наиболее интенсивно кровоснабжаемым органам — ежеминутно через почку проходит */4 всего объема крови, выбрасываемой сердцем, при этом объем кровотока в коре почки, где происходит фильтрация плазмы крови и образование первичной мочи, составляет свыше 90 % общего почечного кровотока. Основной структурно-функциональной единицей почки, обеспечивающей образование мочи, является нефрон. В почке человека находится около 1,2 млн. нефронов. Однако не все нефроны функционируют в почке одновременно, существует определенная периодичность активности отдельных нефронов, когда часть из них функционирует, а другие нет. Эта периодичность обеспечивает надежность деятельности почки за счет функционального дублирования. В связи с этим важным показателем функциональной активности почки является масса действующих нефронов в конкретный момент времени. Нефрон состоит из нескольких последовательно соединенных отделов (рис. 14.2), располагающихся в корковом и мозговом веществе почки. Сосудистый клубочек, или мальпигиево тельце, является структурой, где происходит процесс ультрафильтрации плазмы крови через фильтрационный барьер и образование первичной мочи (рис. 14.3). Он расположен в корковом веществе, имеет около 50 капиллярных петель, связанных друг с другом и подвешенных как на брыжейке с помощью мезангия, состоящего из волокнистых структур и мезангиальных клеток. Снаружи клубочки покрыты двухслойной капсулой Боумена—Шумлянского. Висцеральный листок этой капсулы покрывает капилляры клубочка и состоит из эпителиальных отростчатых клеток — подоцитов. Отростки подоцитов (большие и малые), называемые педикулами, покрывают всю поверхность капилляров, тесно переплетаясь друг с другом и оставляя межпедикулярные пространства не более 30 нм. Пространства заполнены фибриллярными структурами, образующими щелевую диафрагму, формирующую решетку или сито с диаметром пор около 10 нм. Наружный или париетальный листок капсулы состоит из базальной мембраны, покрытой кубическими эпителиальными клетками, переходящими в эпителий канальцев. Между двумя листками капсулы, расположенными наподобие чаши, имеется щель или полость капсулы, в которую происходит ультрафильтрация плазмы крови. Полость капсулы переходит в просвет главного или проксимального отдела канальцев. Стенка всех канальцев нефрона, где по мере продвижения мочи происходит обратное всасывание в кровь воды и различных веществ (реабсорбция), а также секреция веществ из крови в мочу, состоит из эпителиальных клеток, расположенных на базальной мембране (рис. 14.4). По строению и функции у этих клеток выделяют апикальную, или люминальную, мембрану, обращенную в просвет канальца, и базолатеральную мембрану. Канальцевый аппарат нефрона подразделяют на несколько отделов. Главный, или проксимальный, отдел канальцев, начинающийся от полости капсулы извитой частью, которая затем переходит в прямую часть канальца. Клетки проксимального отдела на апикальной мембране имеют щеточную каемку из микроворсин, покрытых гликокаликсом. Проксимальный отдел расположен в корковом веществе, где переходит в петлю Генле. Тонкий нисходящий отдел петли Генле, который покрыт плоскими клетками с щелевидными пространствами в цитоплазме шириной до 7 нм,
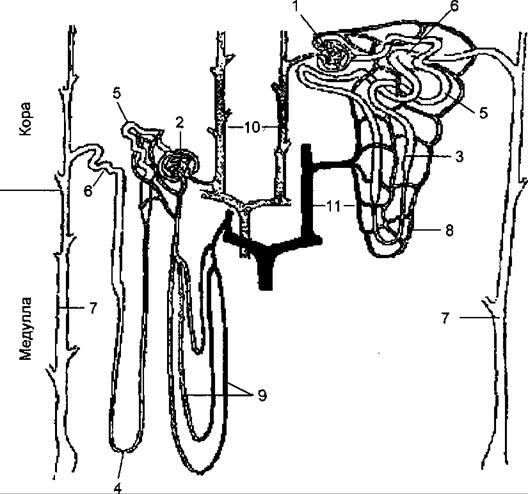
Рис. 14.2. Типы и структура нефронов.
1 — клубочек интракортикального нефрона; 2 — клубочек юкстамедуллярного нефрона; 3 — петля Генле интракортикального нефрона; 4 — петля Генле юкстамедуллярного нефрона; 5 — проксимальные извитые канальцы; 6 —дистальные извитые канальцы; 7 — собирательные трубочки; 8 — капиллярная сеть интракортикального нефрона; 9 — прямые капиллярные сосуды юкстамедуллярного нефрона; 10 — артерии и артериолы; 11 — венулы и вены. Интракортикальные нефроны имеют короткие петли Генле, выносящая артериола клубочка образует густую капиллярную сеть вокруг канальцев. Юкстамедуллярные нефроны имеют длинные петли Генле, спускающиеся вглубь мозгового вещества к почечному сосочку и образующие канальцевую противоточную систему почки, а выносящие артериолы клубочка формируют в мозговом веществе почки прямые нисходящие и восходящие капиллярные сосуды, образующие сосудистую противоточную систему. спускающийся в мозговое вещество почки, где поворачивает на 180° и переходит в тонкую восходящую часть, соединяющуюся с дистальным отделом канальцев. Дистальный отдел канальцев, состоящий из толстой восходящей части петли Генле или прямого отдела и извитой части. Восходящая часть покрыта клетками, напоминающими клетки проксимального отдела, но лишенными щеточной каемки. Восходящая часть дистального канальца вновь входит в кору почки, подходит к клубочку и обязательно соприкасается с его полюсом между приносящей и выносящей артериолами. Здесь эпителий канальца становится цилиндрическим, ядра клеток — гипер- хромными, этот участок выглядит темным, плотным, что и дало ему название macula densa — плотное пятно. Поскольку непрерывная базальная мембрана здесь отсутствует и клетки эпителия канальца имеют тесный
Рис. 14.3. Схема строения клубочка. А — схематическое изображение клубочка в целом, Б — фрагмент трехслойного фильтрационного барьера, В — увеличенный участок фильтрационного барьера. Отчетливо выявляются три слоя барьера: эндотелий капилляра клубочка, базальная мембрана и клетки висцерального листка капсулы Боумена—Шумлянского (подоциты). Фильтрация воды с растворенными в ней веществами происходит из плазмы крови капилляра клубочка через фенестры эндотелия, поры базальной мембраны и щелевые диафрагмы между ножками подоцитов. Все эти структуры фильтрационного барьера имеют отрицательный заряд.
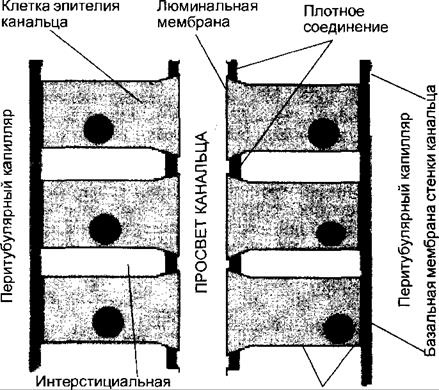
Из просвета канальца через люминальную мембрану клеток эпителия канальца происходит процесс реабсорбции воды и веществ, содержащихся в первичной моче. Поступившие в клетки вещества далее транспортируются через базолатеральные мембраны. Выведение через базолатеральные мембраны натрия в интерстициальную жидкость способствует повышению ее осмотического давления и обеспечивает осмотический транспорт воды. Из интерстициальной жидкости вода и вещества всасываются в кровь перитубулярных капилляров. Клетки эпителия канальцев обладают также способностью захватывать вещества из кровеносных перитубулярных капилляров и затем секретировать их в просвет канальца через люминальные мембраны. контакт с гранулированными миоэпителиоидными клетками артериолы клубочка (юкстагломерулярными клетками), плотное пятно является структурой юкстагломерулярного аппарата почки. Дистальные извитые канальцы через короткий связующий отдел впадают в коре почек в следующий отдел нефрона — собирательные трубки. Собирательные трубки спускаются из коры почек вглубь мозгового вещества, где их эпителий из кубического становится цилиндрическим. Темные цилиндрические эпителиальные клетки дистальных отделов собирательных трубок богаты карбоангидразой и обеспечивают секрецию ионов водорода. В глубине мозгового вещества в области вершин пирамид собирательные трубки сливаются в выводные протоки, открывающиеся в полость лоханки. По особенностям локализации клубочков в коре почек, строения канальцев и особенностям кровоснабжения различают три типа нефронов: суперфициальные, интракортикалъные и юкстамедуллярные (см. рис. 14.2). Суперфициальные нефроны имеют поверхностно расположенные в коре клубочки, наиболее короткую петлю Генле, их 20—30 %. Интракорти- кальные нефроны, клубочки которых расположены в средней части коры почки, наиболее многочисленны (60—70 %) и выполняют основную роль в процессах ультрафильтрации мочи. Диаметр их приносящей артериолы больше, чем у выносящей, ветви последней дают густую сеть капилляров в корковом и мозговом веществе. Юкстамедуллярных нефронов значительно меньше (10—15 %), клубочки их расположены у границы коркового и мозгового вещества почки, выносящие артериолы шире приносящих, петли Генле самые длинные и спускаются почти до вершины сосочка пирамид. Выносящие артериолы образуют прямые капиллярные нисходящие и восходящие сосуды, идущие в глубину мозгового вещества параллельно петлям Генле. Юкстамедуллярные нефроны играют ведущую роль в процессах концентрирования и разведения мочи. Механизм мочеобразования складывается из трех основных процессов: • клубочковой ультрафильтрации из плазмы крови воды и низкомолекулярных компонентов с образованием первичной мочи; • канальцевой реабсорбции (обратного всасывания в кровь) воды и необходимых для организма веществ из первичной мочи; • канальцевой секреции ионов, органических веществ эндогенной и экзогенной природы. 14.2.1.1. Клубочковая ультрафильтрация и ее регуляция Процесс клубочковой ультрафильтрации (далее просто фильтрация) осуществляется под влиянием физико-химических и биологических факторов через структуры гломерулярного фильтра, находящегося на пути выхода жидкости из просвета капилляров клубочка в полость капсулы Боумена— Шумлянского. Гломерулярный фильтр состоит из 3 слоев: эндотелия капилляров, базальной мембраны и эпителия висцерального листка капсулы или подоцитов (см. рис. 14.3). Эндотелий капилляров пронизан отверстиями диаметром до 100 нм. На поверхности эндотелия находится особая выстилка отрицательно заряженными молекулами гликопротеинов, мешающая доступу форменных элементов и крупных молекул, в том числе и белков, к лежащей под эндотелием базальной мембране. Базальная мембрана является основной частью фильтра, препятствующей проникновению из плазмы крови крупномолекулярных соединений (белков). При этом не только размер пор мембраны (около 2,9 нм), но и их отрицательный заряд противодействуют прохождению молекул с отрицательным зарядом, например альбуминов. Базальная мембрана довольно быстро «изнашивается» за счет непрерывного процесса фильтрации, и ее элементы постоянно восстанавливаются с помощью мезангиальных клеток, при этом в течение года происходит полная замена ее основного вещества. Третий слой фильтра образован отростками подоцитов, между которыми остаются щелевые диафрагмы с диаметром пор около 10 нм, поры покрыты гликокаликсом, оставляющим отверстия радиусом около 3 нм. Эта часть фильтра также несет отрицательный заряд. Поскольку подоциты содержат внутри отростков — педикул актомиозиновые миофибриллы, они могут сокращаться и расслабляться, действуя как микронасосы, откачивающие фильтрат в полость капсулы. Эта активность подоцитов составляет один из биологических факторов обеспечения процесса фильтрации, к числу которых относится также сокращение и расслабление мезангиальных клеток, изменяющих тем самым площадь поверхности клубочкового фильтра. Физико-химические факторы обеспечения фильтрации представлены отрицательным зарядом структур фильтра и фильтрационным давлением, являющимся основной причиной фильтрационного процесса. Фильтрационное давление — это сила, обеспечивающая движение жидко-


Приносящая артериола
При сужении выносящей артериолы гидростатическое давление растет и скорость клубочковой фильтрации (СКФ) повышается, а при сужении приносящей артериолы гидростатическое давление и СКФ падают. сти с растворенными в ней веществами из плазмы крови капилляров клубочка в просвет капсулы. Эта сила создается гидростатическим давлением крови в капилляре клубочка. Препятствующими фильтрации силами являются онкотическое давление белков плазмы крови (так как белки почти не проходят через фильтр) и давление жидкости (первичной мочи) в полости капсулы клубочка. Таким образом, фильтрационное давление (ФД) представляет собой разность между гидростатическим давлением крови в капиллярах (Рг) и суммой онкотического давления плазмы крови (Ро) и давления первичной мочи (Рм) в капсуле: ФД - Рг — (Ро + Рм). По ходу капилляров клубочка от приносящего к выносящему отделу гидростатическое давление снижается за счет сосудистого сопротивления, а онкотическое давление плазмы, благодаря потере фильтрующейся воды и сгущению, возрастает. Гидростатическое давление крови в приносящей части капилляров клубочка высокое, примерно 50—60 мм рт. ст., т. е. выше, чем в капиллярах других тканей. Это связано, во-первых, с тем, что капилляры клубочка находятся близко к аорте (короткие почечные и внутрипочечные артерии), и, во-вторых,—диаметр приносящих артериол клубочка больше, чем у выносящих. Гидростатическое давление в выносящей части капилляров ниже на 2—5 мм рт. ст. Гидростатическое давление увеличивается или снижается при изменении соотношения диаметров приносящей и выносящей артериол, что является ведущим механизмом регуляции процесса фильтрации (рис. 14.5). Онкотическое давление белков плазмы крови в приносящей части капилляров клубочка около 25 мм рт. ст., а в выносящей части капилляров, благодаря фильтрации из плазмы воды, оно возрастает до 35—40 мм рт.ст. Давление первичной мочи в капсуле Боумена— Шумлянского примерно равно 15—20 мм рт. ст. Таким образом, ФД в приносящей части капилляров клубочка составляет в среднем: 60 — (25 + 15) = 20 мм рт. ст. В выносящей части капилляров фильтрации практически не происходит, так как ФД равно: 58 — (40 + 15) = 3 мм рт. ст. Основной количественной характеристикой процесса фильтрации является скорость клубочковой фильтрации (СКФ). СКФ — это объем ультрафильтрата или первичной мочи, образующийся в почках за единицу времени. Эта величина зависит от нескольких факторов: 1) от объема крови, точнее плазмы, проходящей через корковое вещество почек в единицу времени, т. е. почечного плазмотока, составляющего в среднем у здорового человека массой 70 кг около 600 мл/мин; 2) фильтрационного давления, обеспечивающего сам процесс фильтрации; 3) фильтрационной поверхности, которая равна примерно 2—3 % от общей поверхности капилляров клубочка (1,6 м2) и может изменяться при сокращении подоцитов и мезангиальных клеток; 4) массы действующих нефронов, т. е. числа клубочков, осуществляющих процесс фильтрации в определенный промежуток времени. В физиологических условиях СКФ поддерживается на довольно постоянном уровне (несмотря на изменения системного артериального давления) за счет механизмов ауторегуляции. К их числу относятся: 1) миоген- ная ауторегуляция тонуса приносящих артериол по принципу феномена Остроумова—Бейлиса; 2) канальцево-клубочковая обратная связь, приводящая к изменению соотношения тонуса приносящих и выносящих артериол клубочка. Вовлечение в регуляцию механизма обратной связи обусловлено изменением доставки с фильтратом в область плотного пятна (macula densa) ионов натрия и хлора, что ведет к изменению продукции в ЮГА гуморальных регуляторов: аденозина (суживающий афферентные артериолы фактор), NO (дилатирующий артериолы фактор), ренина и ангио- тензина-П, кининов и простагландинов (рис. 14.6); 3) изменения числа функционирующих нефронов. Первые два механизма поддерживают постоянство кровотока в клубочках и фильтрационное давление, гуморальные регуляторы могут менять площадь фильтрационной поверхности и функции подоцитов, третий механизм определяет конечный эффект ауторегуляции СКФ в органе, что в итоге обеспечивает постоянство объема образуемой первичной мочи. СКФ определяют в результате сопоставления концентрации определенного вещества в плазме крови и моче. При этом используемое вещество должно выделяться вместе с водой только путем фильтрации и не всасываться в нефроне обратно в кровь. Таким условиям больше всего соответствует полисахарид фруктозы инулин. Исходя из концентрации инулина в плазме [Пин], и, определив его концентрацию в определенном объеме (V) конечной мочи [Мин], рассчитывают, какой объем первичной мочи соответствует найденной концентрации инулина. Насколько выросла концентрация инулина в конечной моче по сравнению с его концентрацией в плазме, во столько раз больше объем профильтровавшейся плазмы (т. е. первичной мочи) объема конечной мочи. Этот показатель получил название «клиренса» инулина или коэффициента очищения и рассчитывается по формуле:

Увеличение загрузки канальцев натрием и хлоридом Увеличение реабсорбции натрия и хлорида клетками - macula densa
Рис. 14.6. Канальцево-клубочковая обратная связь как механизм ауторегуляции клубочковой фильтрации. Сплошные стрелки указывают последовательность реакций, штриховые стрелки ■—участки нефрона, где происходит соответствующий процесс. В качестве примера приведен случай первичного повышения гидростатического давления в капиллярах клубочка при расширении приносящей артериолы. В результате повышенного гидростатического давления растут скорость клубочковой фильтрации, объем образующейся первичной мочи и скорость ее продвижения по канальцам. При повышенной скорости тока мочи в проксимальных канальцах не успевает реабсорбироваться должная часть профильтровавшихся натрия и хлорида, соответственно, увеличивается загрузка дистальных отделов канальцев натрием и хлоридом и их реабсорбция клетками macula densa. Последние увеличивают синтез и секрецию в кровь аденозина, побуждают юкстагломерулярные клетки к активации синтеза и секреции ренина. Поступающий в кровь аденозин, равно как и образующийся в ней под влиянием ренина ангиотензин, вызывают сужение приносящей артериолы клубочка, снижение гидростатического давления и восстановление скорости клубочковой фильтрации. вается обратно в кровь и концентрация инулина растет, что и находят в конечной моче. Поскольку инулин в организме отсутствует, для определения СКФ его необходимо капельно вводить в кровоток, создавая постоянную концентрацию. Это затрудняет исследование, поэтому в клинике обычно используют эндогенное вещество креатинин, концентрация которого в крови довольно стабильна. Таблица 14.1. Основные гуморальные влияния на процессы мочеобразовання
|