Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Регуляторные функции гормонов эндокринных тканей в органах, обладающих неэндокринными функциямиСодержание книги
Поиск на нашем сайте
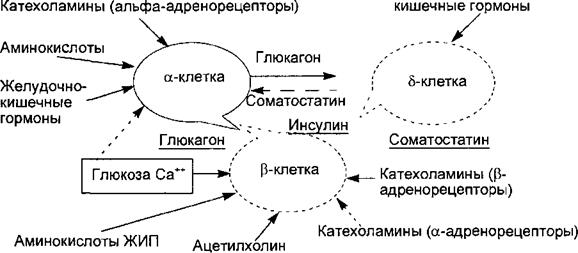
6.7.1. Регуляторные функции гормонов поджелудочной железы Эндокринную функцию в поджелудочной железе выполняют скопления клеток эпителиального происхождения, получившие название островков Лангерганса и составляющие всего 1 —2 % массы поджелудочной железы — экзокринного органа, образующего панкреатический пищеварительный сок. Количество островков в железе взрослого человека очень велико и составляет от 200 тысяч до полутора миллионов. В островках различают несколько типов клеток, продуцирующих гормоны: альфа-клетки образуют глюкагон, бета-клетки — инсулин, дельта-клетки — соматостатин, джи- клетки — гастрин и РР- или F-клетки — панкреатический полипептид. Помимо инсулина в бета-клетках синтезируется гормон амилин, обладающий противоположными инсулину эффектами. Кровоснабжение островков более интенсивно, чем основной паренхимы железы. Иннервация осуществляется постганлионарными симпатическими и парасимпатическими нервами, причем среди клеток островков расположены нервные клетки, образующие нейроинсулярные комплексы. Инсулин синтезируется в эндоплазматическом ретикулуме бета-клеток вначале в виде пре-проинсулина, затем от него отщепляется 23-аминокис- лотная цепь и остающаяся молекула носит название проинсулина. В комплексе Гольджи проинсулин упаковывается в гранулы, в них осуществляется расщепление проинсулина на инсулин и соединительный пептид (С- пептид). В гранулах инсулин депонируется в виде полимера и частично в комплексе с цинком. Количество депонированного в гранулах инсулина почти в 10 раз превышает суточную потребность в гормоне. Секреция инсулина происходит путем экзоцитоза гранул, при этом в кровь поступает
эквимолярное количество инсулина и С-пептида. Определение содержания последнего в крови является важным диагностическим тестом оценки секреторной способности р-клеток. Секреция инсулина является кальцийзависимым процессом. Под влиянием стимула — повышенного уровня глюкозы в крови — мембрана бета- клеток деполяризуется, ионы кальция входят в клетки, что запускает процесс сокращения внутриклеточной микротубулярной системы и перемещение гранул к плазматической мембране с последующим их экзоцитозом.
Секреторная функция разных клеток островков взаимосвязана, зависит от эффектов образуемых ими гормонов, в связи с чем островки рассматриваются как своеобразный «мини-орган» (рис. 6.21). Выделяют два вида секреции инсулина: базальную и стимулированную. Базальная секреция инсулина осуществляется постоянно, даже при голодании и уровне глюкозы крови ниже 4 ммоль/л. Стимулированная секреция инсулина представляет собой ответ бета- клеток островков на повышенный уровень D-глюкозы в притекающей к бета-клеткам крови. Под влиянием глюкозы активируется энергетический рецептор бета-клеток, что увеличивает транспорт в клетку ионов кальция, активирует аденилатциклазу и пул (фонд) цАМФ. Через эти посредники глюкоза стимулирует выброс инсулина в кровь из специфических секреторных гранул. Усиливает ответ бета-клеток на действие глюкозы гормон двенадцатиперстной кишки — желудочный ингибиторный пептид (ЖИП). В регуляции секреции инсулина определенную роль играет и вегетативная нервная система. Блуждающий нерв и ацетилхолин стимулируют секрецию инсулина, а симпатические нервы и норадреналин через альфа-адренорецепторы подавляют секрецию инсулина и стимулируют выброс глюкагона. Специфическим ингибитором продукции инсулина является гормон дельта-клеток островков —- соматостатин. Этот гормон образуется и в кишечнике, где тормозит всасывание глюкозы и тем с; мым уменьшает ответную реакцию бета-клеток на глюкозный стимул. Образование в поджелудочной железе и кишечнике пептидов, аналогичных мостовым, например соматостатина, подтверждает существование в организк е единой APUD-системы. Секреция глюкагона стимулируется снижением уровня глюкозы в крови, гормонами желудочно-кишечного тракта (ЖИП гастрин, секретин, холецистокинин-панкреозимин) и при уменьшении в крови ионов Са2+. Подавляют секрецию глюкагона инсулин, соматостатин, глюкоза крови и Са2+. В эндокринных клетках кишечника образуется глюкагоноподобный пептид-1, стимулирующий всасывание глюкозы и секрецию инсулина после приема пищи. Клетки желудочно-кишечного тракта, продуцирующие гормоны, являются своеобразными «приборами раннего оповещения» клеток панкреатических островков о поступлении пищевых веществ в организм, требующих для утилизации и рас пределе шя участия панкреатических гормонов. Эта функциональная взаимосвязь нашла отражение в термине «гастро-энтеро-панкреатическая система».
6.7.1.1. Физиологические эффекты инсулина Действие инсулина на клетки-мишени начинается после его связывания со специфическими димерными мембранными рецепторами (рис. 6.22), при этом внутриклеточный домен рецептора обладает тирозинкиназной активностью. Инсулин-рецепторный комплекс не только передает сигнал внутрь клетки, но и частично путем эндоцитоза поступает внутрь клетки к лизосомам. Под влиянием лизосомальной протеазы инсулин отщепляется от рецептора, при этом последний либо разрушается, либо возвращается к мембране и вновь встраивается в нее. Многократное перемещение рецептора от мембраны к лизосомам и обратно к мембране носит название рециклизация рецептора. Процесс рециклизации вакен для регуляции колич- ства инсулиновых рецепторов, в частности обеспечения обратной зависимости между концентрацией инсулина и количеством мембранных рецепторов к нему. Образование инсулин-рецепторного комплекса активирует тирозинки- назу, запускающую процессы фосфорилирования внутриклеточных белков. Происходящее при этом аутофосфорилирование рецептора ведет к усилению первичного сигнала. Инсулин-рецепторный комплекс вызывает активирование фосфолипазы С, образование вторичных посредников инозитолтрифосфата и диацилглицерола, активацию протеинкиназы С, ингибирование цАМФ. Участие нескольких систем вторичных посредников объясняет многообразие и различия эффс ктов инсулина в разных тканях. Инсулин оказывает влияние на все виды обмена веществ, способствует анаболическим процессам, увеличивая синтез пикогена, жиров и белков, тормозя эффекты многочисленных контринсулярных гормонов (глюкагона, катехоламинов, глюкокортикоидов и соматотэопина). Все эффекты инсулина по скорости их реализации подразделяют на 4 группы: очень быстрые (через несколько секунд) — гиперполяризация мембран клеток (за исключением гепатоцитов), повышение проницаемэсти для глюкозы, активация Na-K-АТФазы, входа К+ и откачивания Na, подавление Са-насоса и задержка Са2+; быстрые эффекты (в течение нескольких минут) — активация и торможение различных ферментов, подав; яющих катаболизм и усиливающих анаболические процессы; медленные процессы (в течение нескольких часов) — повышенное поглощение аминокислот, изменение син- Внеклеточный домен реце птора Трансмембранный домеь-------- -^-.рецептора Внутриклеточный домен рецептора
Синтез липидов Синтез гликогена Рост и экспрессия Синтез белков Транспорт генов глюкозы Рис. 6.22. Схема механизма х ействия инсулина на клетку-мишень. Взаимодействие инсулина со сгн пифическим рецептором вызывает активацию тирозинкиназы, связанной с внутриклеточный доменом рецептора. Следствием является фосфорилирование внутриклеточных белков, в i ом числе и аутофосфорилирование внутриклеточного домена рецептора инсулина, усиливают;е тирозинкиназную активность. Фосфорилирование внутриклеточных белков ведет к активации ферментов, увеличению синтеза новых белков, в том числе и переносчиков ГЛЮТ-4, обеспечивающих транспорт в клетку молекул глюкозы.
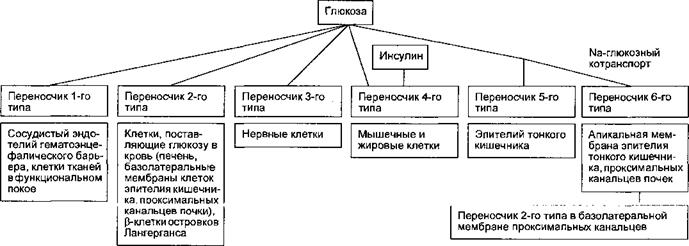
теза РНК и белков-ферме <тов; очень медленные эффекты (от часов до суток) — активация митогенсза и размножения клеток. Важнейшим эффектом инсулина в организме является увеличение в 20—50 раз транспорта тлю <озы через мембраны мышечных и жировых клеток путем облегченной диффузии по градиенту концентрации с помощью чувствительных к гормон) мембранных белковых переносчиков, называемых ГЛЮТ. В мембранах разных видов клеток выявлены 6 типов ГЛЮТ (рис. 6.23), но только один из них — ГЛЮТ-4 — является инсулинозависимым и находится в мембранах клеток скелетных мышц, миокарда, жировой ткани. Инсулин влияет на угле водный обмен, что проявляется: 1) активацией утилизации глюкозы клетками, 2) усилением процессов фосфорилирования; 3) подавлением распаде и стимуляцией синтеза гликогена; 4) угнетением глюконеогенеза; 5) активацией процессов гликолиза; 6) гипогликемией. Действие инсулина на белковый обмен состоит в: 1) повышении проницаемости мембран для ам шокислот; 2) усилении синтеза иРНК; 3) активации в печени синтеза av инокислот; 4) повышении синтеза и подавлении распада белка.
Основные эффекты инс /лина на липидный обмен: • стимуляция синтеза свободных жирных кислот из глюкозы; • стимуляция синтеза липопротеиновой липазы в клетках эндотелия сосудов и благодаря этому активация гидролиза связанных с липопротеинами крови триглицеридов и поступления жирных кислот в клетки жировой ткан1; • стимуляция синтеза триглицеридов; • подавление распада ж лра; • активация окисления кетоновых тел в печени. Благодаря влиянию на } неточную мембрану инсулин поддерживает высокую внутриклеточную концентрацию ионов калия, что необходимо для обеспечения нормальной в эзбудимости клеток. Широкий спектр метаболических эффектов инсулина в организме свидетельствует о том, что гормон необходим для осуществления функционирования всех тканей, оргаьов и физиологических систем, реализации эмоциональных и поведенческих актов, поддержания гомеостазиса, осуществления механизмов приспособления и защиты организма от неблагоприятных факторов среды.
Недостаток инсулина (относительный дефицит по сравнению с уровнем контринсулярных гормонов, прежде всего глюкагона) приводит к сахарному диабету. Избыток инсу; ина в крови, например при передозировке, вызывает гипогликемию с эезкими нарушениями функций центральной нервной системы, использующей глюкозу как основной источник энергии независимо от инсулина. 6.7.7.2. Физиологические эс, мректы глюкагона Глюкагон является мощный контринсулярным гормоном и его эффекты реализуются в тканях череэ систему вторичного посредника аденилатцик- лаза—цАМФ. В отличие о 1 инсулина, глюкагон повышает уровень сахара крови, в связи с чем его называют гипергликемическим гормоном. Основные эффекты глюкагона проявляются в следующих сдвигах метаболизма в организме: • активация гликогенолиза в печени и мышцах; • активация глюконеоп неза; • активация липолиза и подавление синтеза жира в адипоцитах; • повышение синтеза кетоновых тел в печени и угнетение их окисления; • стимуляция катаболи: ма белков в клетках тканей, прежде всего печени, и увеличение синтеза в ней мочевины. Образующиеся в острсвках Лангерганса гастрин и панкреатический полипептид основную рол > играют в регуляции процессов пищеварения, их эффекты и физиолог! ческая роль рассмотрены в соответствующей главе. 6.7.2. Регуляторные функции гормонов половых желез Половые железы (семенники и яичники), наряду с функцией гаметообра- зования, содержат клетки, синтезирующие и секретирующие половые гормоны. При этом эндокринная функция присуща и специализированным для внутренней секреции клеткам (клетки Лейдига семенников, клетки желтого тела яичников), и клеткам, участвующим в процессах гаметогенеза (клетки Сертоли семенников, клетки гранулезы яичников). Как семенники, так и яичники синтезируют и мужские горк оны (андрогены), и женские половые гормоны (эстрогены), являющиеся стероидами ~ производными холестерина. 6.7.2.7. Гормоны семенников и их эффекты в орггнизме Основной структурой семенника, где происходит образование и созревание гамет — сперматозоидов, — являются извит ые семенные канальцы. Базальная мембрана изнутри покрыта отростчатыми клетками Сертоли и располагающимися между ними клетками сгерматогенного эпителия, внутренний слой которых составляют сперматогонии. Клетки Сертоли, наряду с обеспечением процесса созревания сперматид, поглощения остатков их цитоплазмы при превращении сперматиды в сперматозоид, обладают секреторной и инкреторной функцией. Их секреторная функция заключается в выделении в просвет канальца жидкости, в которой плавают сперматозоиды. Инкреторная функция свод1 тся к двум процессам: 1) образованию и секреции с жидкостью в просвет канальца гормона ингиби- на — основного тормозящего продукцию фоллит ропина механизма обратной связи с гипофизом и 2) образованию и секэеции в периканальцевую лимфу эстрогенов. Кровеносные капилляры не проникают в просвет канальцев, а ветвятся между их петлями. Рядом с кровеносными капиллярами расположены скопления клеток мезенхимнсго происхождения, называемых клетками Лейдига. Эти клетки отделены от семенных канальцев лимфатическими пространствами. Клетки Лейдига являются основными продуцентами мужских половых гормонов, главным образом тестостерона. Регуляция продукции гормонов семенниками осуществляется преимущественно лютропином аденогипофиза, специфически регулирующим секреторную активность клеток Лейдига и продукц по тестостерона, и отчасти фоллитропином. меняющим активность клеток Сертоли и продукцию ими эстрогенов и ингибина. Функциональная активность клеток Сертоли и Лейдига регулируется влияниями через лимф} секретируемых ими гормонов.
Основные метаболические и функциональные эффекты тестостерона: • обеспечение процессов половой дифференцировки в эмбриогенезе; • развитие первичных и вторичных половых признаков; • формирование структур центральной нервной системы, обеспечивающих половое поведение и функции; • генерализованное анаболическое действие, обеспечивающее рост скелета, мускулатуры, распределение подкожш го жира; • регуляция сперматогенеза; • задержка в организме азота, калия, фосфата, кальция; • активация синтеза РНК; • стимуляция эритропоэза. 6.7.2.2. Гормоны яичников и их эффекты в организме Гормонопродуцирующие клетки гранулезы фолликулов являются по происхождению и функциям аналогом клеток Сертоли семенников, но их функция регулируется не только гипофизарным фоллитропином, но в большей мере лютропином. Основным гормоном гранулемы является эстрадиол, образуемый из предшественника тестостерона. В меньшем количестве гранулеза образует эстрон, из которого в печени и пл; центе образуется эстриол. Клетки гранулезы образуют в малых количества} и прогестерон, необходимый для овуляции, но главным источником прогестерона служат клетки желтого тела, регулируем ле гипофизарным лютропином. Секреторная активность этих эндокринных клеток характеризуется выраженной цикличностью, связанной с женским половым циклом. Последний обеспечивает интеграцию во времени различных процессов, необходимых для осуществления репродуктивной функции — периодическую подготовку эндометрия к имплантации оплодотворенной яйцеклетки, созревание яйцеклетки и овуляцию, изменение вторичных половых признаков. Эстрогены необходимы для процессов половой дифференцировки в эмбриогенезе, полового созревания и развития женских половых признаков, установления женского полового цикла, роста мышцы и железистого эпителия матки, развития мог очных желез. В итоге эстрогены регулируют половое поведение, овогенез, процессы оплодотворения и имплантации яйцеклетки, развитие и дифференцировку плода, нормальный родовой акт. Благодаря геномному механизму действия эстрогены подавляют резорбцию кости, оказывают общее анаболическое действие, хотя и более слабое, чем андрогены. Негеномгый механизм действия эстрогенов ведет к задержке в организме азота воды и солей. Одним из важных негеномных эффектов эстрогенов является активация под их влиянием NO-синтетазы и образование оксида азе та в коронарных сосудах, что профилактирует развитие у женщин ишемической болезни сердца. Эстрогены модулируют секрецию инсулина и внутриклеточный гомеостазис кальция. Распространенность рецепторов эстрогенов на мембранах разных типов клеток организма (костные, мышечные, секреторные, нервные, соединительной ткани и крови) и, соответственна, множественность эффектов гормонов в организме, объясняют причин г многочисленных функциональных изменений, происходящих в женском организме при климаксе, когда секреция эстрогенов резко снижается. Ввехение в организм женщины в этот период экзогенных веществ с эстрогеноподобным действием, в том числе и растительного происхождения, позголяет существенно уменьшить выраженность и характер функциональных проявлений климактерического периода. Прогестерон является гермоном сохранения беременности (гестагеном), так как ослабляет готовность мускулатуры матки к сокращению. Малые концентрации гормона необходимы и для овуляции. Большие количества прогестерона, образующиеся желтым телом, подавляют секрецию гипофизарных гонадотропинов. Прогестерон обладает выраженным антиальдосте- роновым эффектом, поэтому стимулирует натриурез.
|
|||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2020-12-09; просмотров: 167; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.218.63.176 (0.015 с.) |