Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Регуляторные функции гормонов клеток, сочетающих выработку гормонов и неэндокринные функцииСодержание книги
Поиск на нашем сайте
6.8.1. Регуляторные функции гормонов плаценты Плацента тесно анатомически и функционально связана с организмами матери и плода, поэтому принято говорить о комплексе «мать—плацента- плод», или «фетоплацентаэном комплексе». Синтез в плаценте эстриола происходит не только из эстрадиола матери, но и из дегидроэпиандростерона, образуемого надпочечниками плода. Поэтому по экскреции с мочой матери эстриола можно судить о жизнеспособности плода. В плаценте образуется прогестерон, действующий преимущественно на мускулатуру матки. С плацентарным прогестероном, например, связан временной интервал между рождениями пле дов при двойне. Основная часть гормон эв плаценты у человека по своим свойствам и даже строению напоминает гипофизарные го тадотропин и пролактин. В наибольших количествах при беременности плацентой продуцируется хорионический гонадотропин, оказывающий регул 1торные эффекты не только на процессы дифференцировки и развития п. ода, но и на метаболизм в организме матери. Гормон обеспечивает в орг шизме матери задержку солей и воды, необходимых для растущего плоде, стимулирует секрецию вазопрессина, активирует механизмы иммунитет; у матери. 6.8.2. Регуляторные функции гормонов п имуса Тимус (вилочковая железа) является центральным органом иммунитета, обеспечивающим продукцию специфических Т-лимфоцитов. Тимоциты секретируют в кровь гормональные факторь, оказывающие не только влияние на дифференцировку Т-клеток, но и вызывающие ряд общих регуляторных эффектов в организме. Основные эффекты гормонов тимуса (тимозина, тимопоэтина, тимулина) описаны < главе 8 «Иммунная система». Гормоны тимуса влияют на процессы синтеза клеточных рецепторов к медиаторам и гормонам, стимулируют разруше лие ацетилхолина в нервно- мышечных синапсах, регулируют состояние угг еводного и белкового обмена, а также обмена кальция, функции щитовидной и половых желез, модулируют эффекты глюкокортикоидов, тироксин i (антагонизм) и соматотропина (синергизм). В целом вилочковая железе рассматривается как орган интеграции иммунной и эндокринной систем с рганизма. 6.8.3. Регуляторные функции гормонов п)чек В почках отсутствует специализированная эндокринная ткань, однако ряд клеток обладает способностью к синтезу и сек эеции многих биологически активных веществ, обладающих всеми свойствами классических гормонов. Установленными гормонами почек являются: 1) кальцитриол — третий кальцийрегулирующий гормон, 2) ренин — на1 альное звено ренин-ангио- тензин-альдостероновой системы, 3) эритропоэтин.
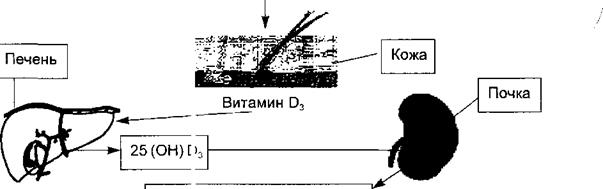
6.8.3.1. Синтез, секреция и физиологические эффекты кальцитриола Кальцитриол является активным метаболитом витамина D3 и в отличие от двух других кальцийрегулирующих гормонов - паратирина и кальцитонина—имеет стероидную природу. Синтез каль щтриола происходит в три этапа (рис. 6.24). Первый этап протекает в кож?, где под влиянием ультрафиолетовых лучей из провитамина образуется штамин D3 или холекалъци- ферол. Второй — связан с печенью, куда холека льциферол транспортируется кровью и где в эндоплазматическом ретикул 'ме гепатоцитов происходит его гидроксилирование по 25-му атому углерод i с образованием 25(OH)D3. Этот метаболит поступает в кровь и циркулирует в связи с альфа-глобулй- ном. Его физиологические концентрации не влияют на обмен кальция. Третий этап осуществляется в почках, где в мттохондриях клеток проксимальных канальцев происходит второе гидроксилирование и образуются два соединения: 1,25-(OH)2-D и 24,25-(OH)2-D. Первое — является наиболее активной формой витамина D3, обладает мощным регуляторным влиянием на обмен кальция в организме и называется кальцитриолом. Образование в почках этого гормона регулируется паратирином, который стимулирует гидроксилирование по первому атому углерода. Таким же эффектом обладает и гипокальциемия. При избытке кальция в крови гидрокси-
Инсоляция
1,25 (ЭН)2 О3или кальцитриол 24, 25 (OH)2D3 Рис. 6.24. Схема образования кальцитриола или активной формы витамина D3.
Под воздействием ультрафиолетовы к лучей в коже из холестерина образуется витамин D3; поступая с кровью в печень, он подвс эгается первому гидроксилированию по 25-му атому углерода, затем из печени с кровью noi адает в почки, где подвергается второму гидроксилированию по 1-му атому углерода, что и i едет к образованию дважды гидроксилированного витамина или кальцитриола.
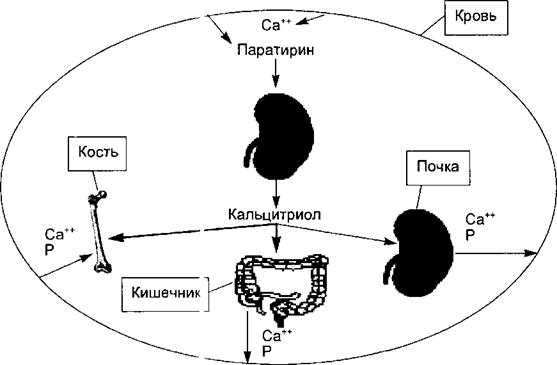
Рис. 6.25. Основные эффекты f альцитриола.
Под влиянием паратирина почка се <ретирует кальцитриол, основные эффекты которого (жирные стрелки) заключаются в стим} пяции всасывания в кишечнике в кровь ионов кальция и фосфата и усилении их захвата кост ной тканью. Эффекты кальцитриола на почку (стимуляция реабсорбции кальция и фосфора) в сражены слабее.
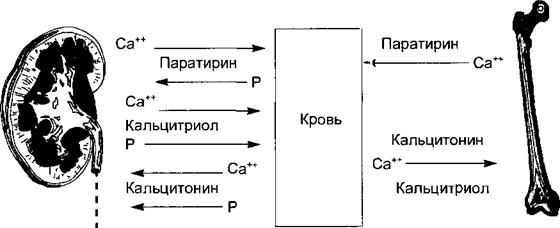
Рис. 6.26. Эффекты трех кальцийрегулирующих горм >нов на органы-мишени. В почке паратирин и кальцитриол активируют реабсорбции кальция, а кальцитонин ее угнетает. И паратирин, и кальцитонин подавляют реабсорбцию < юсфата. В кишечнике кальцитриол и паратирин активируют всасывание кальция и фосфата Кальцитонин и кальцитриол способствуют отложению кальция в костях, а паратирин актп шрует резорбцию кости и выход кальция в кровь. лирование происходит по 24-му атому углерода и синтезируется второе соединение — 24,25-(OH)2-D, которое обладает си >собностью угнетать секрецию паратирина по принципу обратной связи. Инактивация кальцитриола происходит в печени. Основной эффект кальцитриола (рис. 6.25) заключается в активации всасывания кальция в кишечнике. Гормон стих улирует все три этапа всасывания: захват ворсинчатой поверхностью клетки, внутриклеточный транспорт, выброс кальция через базолатеральн 'Ю мембрану во внеклеточную среду. Действие кальцитриола на эпителиальные клетки кишечника состоит в индуцировании синтеза энтероцитаии специальных кальций- связывающих и транспортирующих белков — к алъбайндинов, Кальцитриол повышает в кишечнике и всасывание фосфато и Почечные эффекты гормона заключаются в стимуляции реабсорбции оосфата и кальция канальцевым эпителием. Эффекты кальцитриола на костную ткань связаны с прямой стимуляцией остеобластов и обеспечением костной ткани усиленно всасывающимся в кишечнике кальцием, что активирует рост и минерализацию кости. Эффекты кальцитриола, как и зсех стероидных гормонов, делятся на геномные и негеномные. Геномные эффекты обусловливают синтез кальбайндинов, активирование остеобла< тов и синтеза костной ткани. Увеличение транспорта кальция внутрь клегок, например в скелетных и сердечной мышцах, в остеобластах, энтероцгтах, гепатоцитах и клетках околощитовидных желез, происходит быстро и обусловлено негеномным действием гормона. Под влиянием гормонрецепторного мембранного комплекса в клетках происх< щит образование вторичного посредника диацил- глицерола и активация протеинкиназы С. Кальцитриол меняет в клетке и уровень цАМФ и цГМФ, что ведет к модификации геномного эффекта. Наличие специфических рецепторов к гормону у многих клеток тканей (в молочной железе, эндокринных железах, нервной системе), способность кальцитриола активиров; ть транспорт кальция в большинстве из них свидетельствуют о широком спектре эффектов этого гормона. Участие трех кальцийрегулирующих гормонов в гомеостазисе кальция и фосфора показано на рис. 6.26. Недостаточность кальцитриола проявляется в виде рахита, т. е. нарушения созревания и кальцификации хрящей и кости у детей, либо остеомаляции, т. е. падения минер ишзации костей после завершения роста скелета. При этом сдвиги уровня кальция в крови и клетках обусловливают угнетение нейромышечной возбудимости и мышечную слабость. 6.8.3.2. Образование ренина и основные функции ренин-ангиотензи н-алъдостероновой системы
Ренин образуется в виде г роренина и секретируется в юкстагломерулярном аппарате (ЮГА) (от латинских слов juxta — около, glomerulus — клубочек) почек миоэпителиоидны ии клетками приносящей артериолы клубочка, получившими название юкстагломерулярных (ЮГК). Структура ЮГА приведена на рис. 6.27. В ЮГА кроме ЮГК также входит прилегающая к приносящим артериолам час ь дистального канальца нефрона, многослойный эпителий которого образ} ет здесь плотное пятно — macula densa. Секреция ренина в ЮГК регулируется четырьмя основными влияниями. Во-первых, величиной давления крови в приносящей артериоле, т. е. степенью ее растяжения. Снижение растяжения активирует, а увеличение — подавляет секрецию ренина. Во-втоэых, регуляция секреции ренина зависит от концентрации натрия в моче дистального канальца, которая воспринимается macula densa — своеобразным Na-рецептором. Чем больше натрия оказывается в моче дистального канальца, тем выше уровень секреции ренина. В-третьих, секреция ренина регулируется симпатическими нервами, ветви которых заканчиваются на ЮГК, медиатор норадреналин через бета-адренорецепторы стимулирует секрецию ренина. В-четвертых, регуляция секреции ренина осуществляется по механизму отрицательной обратной связи, включаемой уровнем I крови других компонентов системы — ангиотензина и альдостерона, а т юсе их эффектами — содержанием в крови натрия, калия, артериальным давлением, концентрацией простагландинов в почке, образующихся под влиянием ангиотензина. Кроме почек образование ренина происходит в эндотелии кровеносных сосудов многих тканей, м гокарде, головном мозге, слюнных железах, клубочковой зоне коры надпочечников. Секретированный в кр >вь ренин вызывает расщепление альфа-глобули- на плазмы крови — ангиотензиногена, образующегося в печени. При этом в крови образуется (рис. 6.'.’.8) малоактивный декапептид ангиотензин-1, который в сосудах почек, легких и других тканей подвергается действию превращающего фермента (карбоксикатепсин, кининаза-2), отщепляющего от ангиотензина-1 две аминокислоты. Образующийся октапептид ангиотензин-11 обладает большим числом различных физиологических эффектов, в том числе стимуляцией К1убочковой зоны коры надпочечников, секрети-

импатическая активация I —► ЮГК —Macula —NaCI —— densa Ренин Ангиотензиноген 1 2 3 4 5 6 7 8 < 10 V11 12 13 14 Asp-Arg-Val-Tyr-lle-His-Pro-Phe-Hs-Leu-Leu-Val-Tyr-Ser-R Ангиотензин-I 1 23 4567 8 9 10
Asp-Arg-VakTyr-lle-His-Pro-Phethbs-Leu I— Дипептидкарбоксилаза (ПФ) Ангиотензин-Ц 1 234567 8 Asp-Arg-Val-Tyr-lle-His-Pro-Phe Рис. 6.28. Активация секреции ренина и образование в крови ангиотензина-II. Показаны три вида стимулов для секреции ренина юкст; гломерулярными клетками почек: снижение АД в приносящей артериоле клубочка, повыи ение симпатической активности, влияния macula densa, вызванные сдвигами уровня натрия. Тод влиянием фермента ренина от молекулы белка ангиотензиногена отщепляется декапеп’ид— ангиотензин-I. Этот пептид подвергается воздействию превращающегося фермента (ПФ) дипептидкарбоксилазы клеток эндотелия сосудов легких, почек и др., отщепляющей две инокислоты. Образующийся октапептид является ангиотензином-П, • вызывает сужение артериальных сосудов, • активирует симпатиче* кую нервную систему как на уровне центров, так и способствуя сип гезу и освобождению норадреналина в синапсах, • повышает сократимое! ь миокарда, • увеличивает реабсорбтию натрия и ослабляет клубочковую фильтрацию в почках, • способствует формиро;анию чувства жажды и питьевого поведения. Таким образом, ренин-а!гиотензин-альдостероновая система участвует в регуляции системного и почечного кровообращения, объема циркулирующей крови, водно-солев зго обмена и поведения. 6.8.4. Регуляторные эфректы гормонов сердца Миоцитами предсердий (преимущественно правого) образуется пептидный гормон с установленной химической структурой, получивший название предсердный натрийуретический гормон, или атриопептид. Гормон накапливается в специфических i ранулах саркоплазмы миоцитов и секретируется в кровь под влиянием гида регуляторных стимулов: растяжения предсердий объемом крови, урозня натрия в крови, эффектов блуждающего и симпатических нервов, содержания в крови вазопрессина. Физиологические эффе] гы атриопептида многообразны, так как во многих органах и тканях обнаружены специфические для него мембранные рецепторы (рис. 6.29). Сосудистые эффекты гормона состоят в расслаблении гладких мышц с юудов и вазодилатации (через цАМФ), снижении артериального давления. Кроме того, гормон повышает проницаемость гистогематических барьеров и увеличивает транспорт воды из крови в тканевую жидкость.
    
Почечные эффекты атриопептида включают: 1) повышение экскреции натрия (до 90 раз) и хлора (до 50 раз) в связи с подавлением их реабсорбции в канальцах. Гормон — в 1000 раз более эффективный натрийуретик, чем фуросемид; 2) выраженное диуретическое действие за счет увеличения клубочковой фильтрации и подавления реабсорбции воды; 3) подавление секреции ренина, ингибирование эффектов ангиотензина-П и альдостерона, т. е. гормон является полным антагонистом РААС.
Атриопептид, кроме того, расслабляет гладкую мускулатуру кишечника, уменьшает величину внутриглазного давления, объема и давления ликвора в желудочках мозга. Близкий по натрийуретическому эффекту гормон выявляется и в ткани головного мозга. Помимо атриопептида в предсердиях образуются атриопептины, повышающие артериальное давление и обладающие антидиуретическим эффектом. Атриопептины участвуют в регуляции питьевого поведения, солевого аппетита и жажды. В малых количествах в сердце образуются соматостатин, ангиотензин-II и релаксин, обладающие хронотропным влиянием на миокард. 6.8.5. Регуляторная функция гормонов сосудистого эндотелия Клетки сосудистого эндотелия синтезируют и выделяют через апикальную и базальную мембраны три группы гормонов: сосудосуживающие (эндоте- лины, тромбоксаны), сосудорасширяющие (оксид азота, гиперполяризую- щий фактор, простагландины) и факторы адгезии и агрегации клеточных элементов. Эндотелины (ЭТ) являются крупными полипептидами (21 аминокислота), образуются путем частичного гидролиза молекулы предшественника или «большого эндотелина», состоящей из 38 аминокислот, под влиянием связанного с мембраной клетки и находящегося в цитоплазматических везикулах эндотелинпревращающего фермента. Этот фермент локализован в эндотелии сосудов легких, сердца, почек, плаценты, поджелудолчной железы, надпочечников, головного мозга и даже в сосудистых гладких мышцах. Наличие двух форм эндотелинпревращающего фермента (мембраносвязанной и внутриклеточной) определяет и два места образования эндоте- линов — в цитоплазме и на поверхности клеточной мембраны. Эндотелин- превращающий фермент ведет не только к образованию молекул эидоте- лина, но способен вызывать гидролиз и инактивацию на поверхности эн- дотелиоцитов ряда регуляторных пептидов и гормонов (инсулина, брадикинина, нейротензина и др.). Активация эндотелинпревращающего фермента происходит под влиянием цитокинов. Эффекты эндотелинов обусловлены их взаимодействием со специфическими мембранными рецепторами двух типов — ЭТ-А и ЭТ-В. Следствием специфического связывания эндотелина с рецептором является активация систем вторичных посредников (фосфолипаза С, ИФЗ, диацилглицерол, цГМФ и цАМФ, фосфолипазы D и А2). Различают прямые и опосредованные сосудистые эффекты эндотелинов. Прямые эффекты заключаются в действии на гладкие мышцы сосудов. Связывание эндотелина с рецепторами гладких мышц сосудов вызывает их сокращение и вазоконстрикцию (через образование ИФЗ и повышение внутриклеточного Са2+), а также стимуляцию митогенеза и пролиферации клеток (^ерез активацию тиро- зинкиназы и фосфорилирование тирозина). Взаимодействие эндотелинов с рецепторами клеток эндотелия вызывает реализацию опосредованных эффектов, в виде высвобождения из эндотелия вазоактивных факторов, приводящих к расширению сосудов (NO, гиперполяризующий фактор, простагландины). Кроме сосудистых эффектов эндотелины изменяют секрецию гормонов гипофиза и надпочечников, стимулируют секрецию атриопептида миокардом, угнетают эффекты вазопрессина в почках, способствуя диурезу и натрийурезу, увеличивают реакции сердца на симпатические стимулы. Среди сосудорасширяющих гормонов эндотелия основное место по выраженности и распространенности эффекта занимает оксид азота (NO), постоянно образующийся из L-аргинина под влиянием фермента NO-син- тетазы. Одним из стимулов, активирующих фермент и образование оксида азота, является механическое растяжение стенки сосудов. Активация фермента и синтез оксида азота происходят при действии на мембранные рецепторы эндотелиоцитов ацетилхолина, адреномедуллина, гистамина, брадикинина, АТФ, а также в результате повышения в клетке эндотелия концентрации ионизированного Са2+. NO-синтетаза помимо образования оксида азота стимулирует синтез некоторых цитокинов: интерлейкина — 1р, альфа-интерферона, тогда как другие цитокины: ИЛ-4, ИЛ-8, ИЛ-10, напротив, подавляют активацию фермента. Вазодилатирующий эффект NO опосредуется активацией образования в гладкомышечных клетках цГМФ. Оксид азота также подавляет вазоконстрикторное действие ангиотензина-П. Синтезированный клетками эндотелия оксид азота выделяется не только через базальную мембрану в сторону гладкомышечных клеток кровеносных сосудов, но и через апикальную мембрану, где тормозит адгезию тромбоцитов и лейкоцитов крови к эндотелиальной выстилке стенки сосуда. Антиагрегантное действие оксида азота отчасти опосредовано простациклином, образующимся в эндотелии. В нервной системе NO является модулятором синаптической передачи, так как выявлено его поступление в синаптическую щель и показано ингибирующее влияние на выделение медиаторных аминокислот. Эндотелиальный гиперполяризующий фактор также вызывает дилатацию артериальных и венозных сосудов, но отличается от NO механизмом действия — способностью активировать ионные каналы (К+,СГ) и снижать возбудимость эндотелиальных клеток. К числу факторов регуляции адгезии и агрегации клеток относятся многочисленные интегрины и селектины, образуемые клетками эндотелия. 6.8.6. Регуляторная функция гормонов желудочно-кишечного тракта Клетки эпителия желудка и двенадцатиперстной кишки обладают способностью секретировать большое количество пептидных соединений, многие из которых выявляются также в головном мозге. Это дало основание рассматривать продуцирующие пептиды клетки в качестве единой APUD-сис- темы организма. Основной функцией гастроинтестинальных гормонов является регуляция процессов пищеварения, а мозговых пептидов — пищевого поведения. В связи с этим гастроинтестинальные гормоны, их природа и регуляторные эффекты представлены в главе 15 «Функции пищеварительной системы».
|
||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2020-12-09; просмотров: 111; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.45.223 (0.016 с.) |